The Genetics of Equine Coat Color
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

I . the Color Gene C
THE ABC OF COLOR INHERITANCE IN HORSES W. E. CASTLE Division of Genetics, University of California, Berkeley, California Received October, 27, 1947 HE study of color inheritance in horses was begun in the early days of Tgenetics. Indeed many facts concerning it had already been established earlier, by DARWINin his book on “Variation of Animals and Plants under Domestication.” At irregular inteivals since then, new attempts have been made to collect and classify in terms of genetic factors the records contained in stud books concerning the colors of colts in relation to the colors of their sires and dams. A full bibliography is given by CREWand BuCHANAN-SMITH (19301. By such studies, we have acquired very full information as to what color a colt may be expected to have, when the color of its parents and grandparents is known. This knowledge is empirical rather than experimental in nature. For horses being slow breeding and expensive are rarely available for direct experi- mental study, such as can be made with the small laboratory mammals, mice, rats, rabbits and guinea pigs. We have definite information that color inheritance in horses involves the existence of mutant genes similar to those demonstrated by experimental studies to be involved in color inheritance of other mammals. But the horse genes have been given special names, as they were successively discovered, and it is difficult at present to correlate them with the better known names and geneic symbols used by the experimental breeders. The present paper is an attempt to make such a correlation. Just as in morphological studies comparative anatomy was found useful and still is used to establish homologies between systems of organs, so in mammalian genetics, a comparative study of gene action in the production of coat colors and color patterns may also be of value. -

Use of Genomic Tools to Discover the Cause of Champagne Dilution Coat Color in Horses and to Map the Genetic Cause of Extreme Lordosis in American Saddlebred Horses
University of Kentucky UKnowledge Theses and Dissertations--Veterinary Science Veterinary Science 2014 USE OF GENOMIC TOOLS TO DISCOVER THE CAUSE OF CHAMPAGNE DILUTION COAT COLOR IN HORSES AND TO MAP THE GENETIC CAUSE OF EXTREME LORDOSIS IN AMERICAN SADDLEBRED HORSES Deborah G. Cook University of Kentucky, [email protected] Right click to open a feedback form in a new tab to let us know how this document benefits ou.y Recommended Citation Cook, Deborah G., "USE OF GENOMIC TOOLS TO DISCOVER THE CAUSE OF CHAMPAGNE DILUTION COAT COLOR IN HORSES AND TO MAP THE GENETIC CAUSE OF EXTREME LORDOSIS IN AMERICAN SADDLEBRED HORSES" (2014). Theses and Dissertations--Veterinary Science. 15. https://uknowledge.uky.edu/gluck_etds/15 This Doctoral Dissertation is brought to you for free and open access by the Veterinary Science at UKnowledge. It has been accepted for inclusion in Theses and Dissertations--Veterinary Science by an authorized administrator of UKnowledge. For more information, please contact [email protected]. STUDENT AGREEMENT: I represent that my thesis or dissertation and abstract are my original work. Proper attribution has been given to all outside sources. I understand that I am solely responsible for obtaining any needed copyright permissions. I have obtained needed written permission statement(s) from the owner(s) of each third-party copyrighted matter to be included in my work, allowing electronic distribution (if such use is not permitted by the fair use doctrine) which will be submitted to UKnowledge as Additional File. I hereby grant to The University of Kentucky and its agents the irrevocable, non-exclusive, and royalty-free license to archive and make accessible my work in whole or in part in all forms of media, now or hereafter known. -

Official Standard of the French Bulldog General Appearance
Official Standard of the French Bulldog General Appearance: The French Bulldog has the appearance of an active, intelligent, muscular dog of heavy bone, smooth coat, compactly built, and of medium or small structure. Expression alert, curious, and interested. Any alteration other than removal of dewclaws is considered mutilation and is a disqualification. Proportion and Symmetry - All points are well distributed and bear good relation one to the other; no feature being in such prominence from either excess or lack of quality that the animal appears poorly proportioned. Influence of Sex - In comparing specimens of different sex, due allowance is to be made in favor of bitches, which do not bear the characteristics of the breed to the same marked degree as do the dogs. Size, Proportion, Substance: Weight not to exceed 28 pounds; over 28 pounds is a disqualification. Proportion - Distance from withers to ground in good relation to distance from withers to onset of tail, so that animal appears compact, well balanced and in good proportion. Substance - Muscular, heavy bone. Head: Head large and square. Eyes dark in color, wide apart, set low down in the skull, as far from the ears as possible, round in form, of moderate size, neither sunken nor bulging. In lighter colored dogs, lighter colored eyes are acceptable. No haw and no white of the eye showing when looking forward. Ears Known as the bat ear, broad at the base, elongated, with round top, set high on the head but not too close together, and carried erect with the orifice to the front. The leather of the ear fine and soft. -

The Occurrence of Silver Dilution in Horse Coat Colours
ISSN 1392-2130. VETERINARIJA IR ZOOTECHNIKA (Vet Med Zoot). T. 60 (82). 2012 THE OCCURRENCE OF SILVER DILUTION IN HORSE COAT COLOURS Erkki Sild, Sirje Värv and Haldja Viinalass Institute of Veterinary Medicine and Animal Sciences, Estonian University of Life Sciences Kreutzwaldi 1, 51014 Tartu, Estonia; Tel: +372 731 3469; Fax: +372 742 2344; e-mail: [email protected] Abstract. The MC1R allele “e” in the homozygous state leads to the production of pheomelanin and is responsible for inhibition of expression of the silver dilution gene (PMEL17 “Z” allele). Horse coat colour is one of the traits breeders select for. A total of 133 horses representing Estonian Native (48), Estonian Heavy Draught (40) and Tori (45) breeds were genotyped for key polymorphisms at C901T in MC1R, the 11 bp deletion in ASIP and C1457T in PMEL17 to determine horse coat colour variation and selection possibilities to increase silver-diluted colours. Our genotyping results showed the “ee” genotype frequency in the MC1R gene to be as follows: Estonian Native 45.8%, Estonian Heavy Draught 65.0%, and Tori 77.8%, and the “Z_” genotype in PMEL17 to be 10.4%, 12.5%, and 0.0%, respectively. Six of total 133 horses with silver dilution were examined for MCOA. No eye abnormalities were detected. Considering the PMEL17 gene singly, silver coat colour could be expressed phenotypically in 12% of genotyped Estonian Heavy Draught horses, but due to unfavourable covariation with the MC1R “e” allele, it only occurred in two per cent of horses. Keywords: ASIP, horse coat colour, MC1R, MCOA, PMEL17, silver phenotype. -

The Show of Colours
THE SHOW OF COLOURS Celebration of every horse SHOWING CLASSES FOR ALL colour – classes for all WITH EVENING PERFORMANCE SPECTACULAR Sunday 7th July 2019 CLASSES FOR ALL HORSES AND PONIES COLOURED, TWO TONE AND SOLID £13.00 Pre entry £15.00 entry on the day Fun classes £10.00 – pre entry and on the day EVENING PERFORMANCE INCLUDING COLOUR PARADE, CLASS AND COLOUR CHAMPIONSHIPS QUALIFYING SHOW FOR CHAPS (UK) OPPORTUNITY TO GAIN POINTS FOR THE HOWE GRAND FINALE AND STEP TOWARDS THE CHANCE TO WIN £500 FOR YOURSELF AND £500 FOR YOU CHOSEN CHARITY QUALIFYING SHOW FOR OUR NEW ECOSSE ELITE TROPHY FOR SCOTTISH BREEDS THE CHANCE TO WIN £100 FOR YOURSELF AND £100 FOR YOU CHOSEN CHARITY QUALIFYING SHOW FOR THE CALEDONIAN SHOWING CHAMPIONSHIPS Please be aware that “Not Before Times” are only as a guide, classes will not start before these times but may start considerably later depending on entries Pre enter on Equo Pre entries close on 3rd July 2019 THE SHOW OF COLOURS – SUNDAY 7th JULY 2019 Welcome to the schedule of the Show of Colours which run in four rings; Ring 1 - Coloured Ring – for all Skewbald and Piebald Ring 2 -Two Tone Ring – for all Roan, Palomino, Spotted and Dun/Buckskin Ring 3 - Solid Ring – for all Black, Grey, Chestnut and Bay Ring 4 - Fun Ring - for all colours Ring 5 - TGCA Scottish Regional Show - The day of showing classes for all horses and ponies of all breeds and types will culminate in an Evening Performance Spectacular. EVENING PERFORMANCE (not before 4.30pm) CONCOURS DE ELEGANCE COLOUR PARADE Free entry – open to all please take part in hand or ridden, details on next page. -

Quantitative Genetic Analysis of Melanoma and Grey Level in Lipizzan Horses
7th World Congress on Genetics Applied to Livestock Production, August 19-23, 2002, Montpellier, France QUANTITATIVE GENETIC ANALYSIS OF MELANOMA AND GREY LEVEL IN LIPIZZAN HORSES I. Curik1, M. Seltenhammer2 and J. Sölkner3 1Animal Science Department, Faculty of Agriculture, University of Zagreb, Croatia 2Clinic of Surgery and Ophthalmology, University of Veterinary Medicine Vienna, Austria 3Department of Livestock Science, University of Agricultural Sciences Vienna, Austria INTRODUCTION Changes of coat colour in which the "dark (non-grey)" colour present in foals is progressively replaced by grey is a known phenomenon in horses. A similar process is present in humans. The grey coat colour is inherited as a dominant trait and is the characteristic, although not exclusive, colour for some horse breeds (Bowling, 2000). The Lipizzan horse, originally bred for show and parade at the Imperial Court in Vienna, is among those breeds. Unfortunately, melanomas (skin tumours) are more prevalent in grey than in non-grey horses (e.g. Seltenhammer, 2000). The causative relationship for this positive association as well as the molecular basis for both traits (melanoma and grey level) are not known (Rieder, 1999 ; Seltenhammer, 2000). The inheritance of coat colour in horses has been always studied from a qualitative view (Sponenberg, 1996 ; Bowling, 2000). In the present study we quantified the grey level (shade) and estimated the proportion of additive genetic component (heritability in the narrow sense) of this trait. Further, we estimated the genetic relationship between melanoma stages and grey level as well as the additive inheritance of melanoma stages. MATERIAL AND METHODS Horses. Data for this study was collected from 351 grey Lipizzan horses of four national studs (Djakovo – Croatia ; 64 horses, Piber – Austria ; 160 horses, Szilvesvarad – Hungary ; 67 horses and Topol'cianky – Slovakia ; 60 horses). -

Horse Sale Update
Jann Parker Billings Livestock Commission Horse Sales Horse Sale Manager HORSE SALE UPDATE August/September 2021 Summer's #1 Show Headlined by performance and speed bred horses, Billings Livestock’s “August Special Catalog Sale” August 27-28 welcomed 746 head of horses and kicked off Friday afternoon with a UBRC “Pistols and Crystals” tour stop barrel race and full performance preview. All horses were sold on premise at Billings Live- as the top two selling draft crosses brought stock with the ShowCase Sale Session entries $12,500 and $12,000. offered to online buyers as well. Megan Wells, Buffalo, WY earned the The top five horses averaged $19,600. fast time for a BLS Sale Horse at the UBRC Gentle ruled the day Barrel Race aboard her con- and gentle he was, Hip 185 “Ima signment Hip 106 “Doc Two Eyed Invader” a 2009 Billings' Triple” a 2011 AQHA Sorrel AQHA Bay Gelding x Kis Battle Gelding sired by Docs Para- Song x Ki Two Eyed offered Loose Market On dise and out of a Triple Chick by Paul Beckstead, Fairview, bred dam. UT achieved top sale position Full Tilt A consistant 1D/ with a $25,000 sale price. 486 Offered Loose 2D barrel horse, the 16 hand The Beckstead’s had gelding also ran poles, and owned him since he was a foal Top Loose $6,800 sold to Frank Welsh, Junction and the kind, willing, all-around 175 Head at $1,000 or City OH for $18,000. gelding was a finished head, better Affordability lives heel, breakaway horse as well at Billings, too, where 69 head as having been used on barrels, 114 Head at $1,500+ of catalog horses brought be- poles, trails, and on the ranch. -

Homozygous Tobiano and Homozygous Black Could Be Winners for Your Breeding Program, If You Know How to Play Your Cards
By IRENE STAMATELAKYS Homozygous tobiano and homozygous black could be winners for your breeding program, if you know how to play your cards. L L I T S K C O T S N N A Y S E T R U O C n poker, a pair is not much to brag gets one of the pair from the sire and the in equine color genetics. If your goal about. Two pairs are just a hair bet - other of the pair from the dam.” is a black foal, and you’ve drawn the ter. But in equine color genetics, a Every gene has an address—a spe - Agouti allele, you’re out of luck. pair—or, even better, two—could cific site on a specific chromosome. be one of the best hands you’ll ever We call this address a locus—plural The Agouti effect hold. We’re talking about a sure bet— being loci. Quite often, geneticists use Approximately 20 percent of horses a pair of tobiano or black genes. the locus name to refer to a gene. registered with the APHA are bay. If Any Paint breeder will tell you that When a gene comes in different you also include the colors derived producing a quality foal that will forms, those variations are called alle - from bay—buckskin, dun, bay roan bring in top dollar is a gamble. In this les. For example, there is a tobiano and perlino—almost one-quarter of business, there are no guarantees. But allele and a non-tobiano allele. Either registered Paints carry and express the what if you could reduce some of the one can occur at the tobiano locus, Agouti allele, symbolized by an upper - risk in your breeding program as well but each chromosome can only carry case A. -
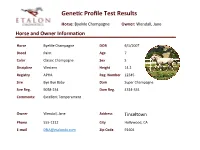
Gene C Profile Test Results
GeneGc Profile Test Results Horse: ByeMe Champagne Owner: Wendall, Jane Horse and Owner Informaon Horse ByeMe Champagne DOB 6/1/2007 Breed Paint Age 7 Color Classic Champagne Sex S Discipline Western Height 14.2 Registry APHA Reg. Number 12345 Sire Bye Bye Baby Dam Super Champagne Sire Reg. 9058-234 Dam Reg. 4314-334 Comments: Excellent Temperament Owner Wendall, Jane Address Tinseltown Phone 555-1212 City Hollywood, CA E-mail [email protected] Zip Code 91604 GeneGc Profile Test Results Horse: ByeMe Champagne Owner: Wendall, Jane Results Summary Coat Color : ByeMe Champagne has one Black allele and one Red allele making the Base coat appear Black. Also detected were single Champagne and Cream alleles; likely resulUng in a rare Champagne Cream color. One copy of the Frame Overo allele is also detected, indicang underlying white patches (hidden By CH). As a result of single gene copies in each of the following, he has a 50% chance of passing Black or Red, Cream and/or Champagne, AgouU and/or Frame Overo alleles to his offspring. Allele Summary: Aa, Ee, Ch, Cr, LWO/n Traits: ByeMe Champagne is a not a carrier of any known recessive disease genes. CauUon is recommended however, as any mare Bred to him should Be Frame Overo negave as to avoid a 25% chance of foal death (+/+ LWO results in a lethal condiUon at Birth). He may also throw Gaited foals when Bred to Gaited (+) mares. Notes: Please note that your analysis is ongoing and may include some regions marked with an asterisk denoUng the following: * Discovery – This gene detecUon is in the -

The Base Colors: Black and Chestnut the Tail, Called “Foal Fringes.”The Lower Legs Can Be So Pale That It Is Let’S Begin with the Base Colors
Foal Color 4.08 3/20/08 2:18 PM Page 44 he safe arrival of a newborn foal is cause for celebration. months the sun bleaches the foal’s birth coat, altering its appear- After checking to make sure all is well with the mare and ance even more. Other environmental issues, such as type and her new addition, the questions start to fly. What gender quality of feed, also can have a profound effect on color. And as we is it? Which traits did the foal get from each parent? And shall see, some colors do change drastically in appearance with Twhat color is it, anyway? Many times this question is not easily age, such as gray and the roany type of sabino. Finally, when the answered unless the breeder has seen many foals, of many colors, foal shed occurs, the new color coming in often looks dramatical- throughout many foaling seasons. In the landmark 1939 movie, ly dark. Is it any wonder that so many foals are registered an incor- “The Wizard of Oz,” MGM used gelatin to dye the “Horse of a rect—and sometimes genetically impossible—color each year? Different Color,” but Mother Nature does a darn good job of cre- So how do you identify your foal’s color? First, let’s keep some ating the same spectacular special effects on her foals! basic rules of genetics in mind. Two chestnuts will only produce The foal’s color from birth to the foal shed (which generally chestnut; horses of the cream, dun, and silver dilutions must have occurs between three and four months of age) can change due to had at least one parent with that particular dilution themselves; many factors, prompting some breeders to describe their foal as and grays must always have one gray parent. -

Busby, D. & Rutland, C. (2019). the Horse. a Natural History. Brighton
Review of: Busby, D. & Rutland, C. (2019). The ed. The origins of the modern domestic horse are Horse. A Natural History. Brighton: Ivy Press. explored in depth, portraying the tarpan as its an- 224 pages, 225 figures (partly colour), hard cover. cestor and Przewalskis as the sole wild horse spe- ISBN 978-1-78240-565-8. cies still in existence. However, this is now outdat- ed knowledge as has been shown by new research, Helene Benkert published in the last two years. GAUNITZ ET AL (2018) and FAGES ET AL (2019) examined and reviewed This richly illustrated book is a well-presented DNA samples of horses from a variety of periods compendium of the horse, its biology, evolution and regions in order to investigate the genetic ori- and its history with humanity. Clearly structured gin of the modern horse. Their research clearly and organised, it is a compelling account of an shows that Przewalski horses are not the last living exceptional species. Throughout, the authors’ re- species of wild horses previously thought, but in gard and respect for horses is apparent but does fact descendants of some of the earliest domesti- not hinder the scientific narrative. On the contra- cated horses. The Eneolithic site of Botai in modern ry, the positive approach to communication, in Kazakhstan yielded some of the earliest evidence combination with plenty of high-quality photo- of horse husbandry and domestication (OUTRAM graphs and schematics, supports the reader’s in- ET AL., 2009). Genetic analysis of the Botai horses terest and inspires to learn more. shows that they are direct ancestors of Przewalski It is largely written in a straightforward and horses but a minimal component in modern do- adequately flowing language, with a clear sen- mestic horses. -

Genetic Test Results
Genetic Profile Test Results HORSE ID: 041418 003 Horse: Merlin PACK: 1 Owner: Alecia Baxley Horse and Owner Information Horse Merlin DOB 2018-04-04 Breed Paint (Overo) Age 0 years, 0 months Color Chestnut/Overo Sex Stallion Discipline Halter Height . Registry APHA Reg Number Pending Sire Ententions Dam Shes Forever Cool Sire Reg & No. Quarter 5605512 Dam Reg & No. APHA 996357 Comments . Owner Alecia Baxley Address 968 County Road 2117 Phone 281 592 6550 City, State Cleveland, TX Email [email protected] Postal Code 77327 [email protected] 650.380.2995 www.EtalonDx.com Genetic Profile Test Results HORSE ID: 041418 003 Horse: Merlin PACK: 1 Owner: Alecia Baxley Results Summary Coat Color: Merlin has two Red alleles and no Black, indicating his base coat color appears Red. One Dominant White 20 allele and one Frame/Lethal White Overo allele was also detected which may result in White markings. As a result of the allele count in each of the following, he has a minimum 100% chance of passing Red, and 50% Dominant White 20 and/or Frame/Lethal White Overo to any offspring. Allele aa, ee, W20/n, LWO/n, HYPP/n, CC (Sprint Type) Summary: Traits: Merlin's testing indicated the presence of one Frame/Lethal White Overo (LWO) allele resulting in “Carrier” status. Caution is recommended when breeding to avoid another carrier and thus, 25% chance of foal death. His testing also indicated the presence of one Hyperkalemic Periodic Paralysis (HYPP) allele indicating “Carrier” and “Possibly Affected” status. Please consult with your veterinarian regarding any medical questions or advice.