How Do Stream Confluences Influence Aquatic Invertebrate Taxonomic and Functional Diversity?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Geomorphic Classification of Rivers
9.36 Geomorphic Classification of Rivers JM Buffington, U.S. Forest Service, Boise, ID, USA DR Montgomery, University of Washington, Seattle, WA, USA Published by Elsevier Inc. 9.36.1 Introduction 730 9.36.2 Purpose of Classification 730 9.36.3 Types of Channel Classification 731 9.36.3.1 Stream Order 731 9.36.3.2 Process Domains 732 9.36.3.3 Channel Pattern 732 9.36.3.4 Channel–Floodplain Interactions 735 9.36.3.5 Bed Material and Mobility 737 9.36.3.6 Channel Units 739 9.36.3.7 Hierarchical Classifications 739 9.36.3.8 Statistical Classifications 745 9.36.4 Use and Compatibility of Channel Classifications 745 9.36.5 The Rise and Fall of Classifications: Why Are Some Channel Classifications More Used Than Others? 747 9.36.6 Future Needs and Directions 753 9.36.6.1 Standardization and Sample Size 753 9.36.6.2 Remote Sensing 754 9.36.7 Conclusion 755 Acknowledgements 756 References 756 Appendix 762 9.36.1 Introduction 9.36.2 Purpose of Classification Over the last several decades, environmental legislation and a A basic tenet in geomorphology is that ‘form implies process.’As growing awareness of historical human disturbance to rivers such, numerous geomorphic classifications have been de- worldwide (Schumm, 1977; Collins et al., 2003; Surian and veloped for landscapes (Davis, 1899), hillslopes (Varnes, 1958), Rinaldi, 2003; Nilsson et al., 2005; Chin, 2006; Walter and and rivers (Section 9.36.3). The form–process paradigm is a Merritts, 2008) have fostered unprecedented collaboration potentially powerful tool for conducting quantitative geo- among scientists, land managers, and stakeholders to better morphic investigations. -

Stream Restoration, a Natural Channel Design
Stream Restoration Prep8AICI by the North Carolina Stream Restonltlon Institute and North Carolina Sea Grant INC STATE UNIVERSITY I North Carolina State University and North Carolina A&T State University commit themselves to positive action to secure equal opportunity regardless of race, color, creed, national origin, religion, sex, age or disability. In addition, the two Universities welcome all persons without regard to sexual orientation. Contents Introduction to Fluvial Processes 1 Stream Assessment and Survey Procedures 2 Rosgen Stream-Classification Systems/ Channel Assessment and Validation Procedures 3 Bankfull Verification and Gage Station Analyses 4 Priority Options for Restoring Incised Streams 5 Reference Reach Survey 6 Design Procedures 7 Structures 8 Vegetation Stabilization and Riparian-Buffer Re-establishment 9 Erosion and Sediment-Control Plan 10 Flood Studies 11 Restoration Evaluation and Monitoring 12 References and Resources 13 Appendices Preface Streams and rivers serve many purposes, including water supply, The authors would like to thank the following people for reviewing wildlife habitat, energy generation, transportation and recreation. the document: A stream is a dynamic, complex system that includes not only Micky Clemmons the active channel but also the floodplain and the vegetation Rockie English, Ph.D. along its edges. A natural stream system remains stable while Chris Estes transporting a wide range of flows and sediment produced in its Angela Jessup, P.E. watershed, maintaining a state of "dynamic equilibrium." When Joseph Mickey changes to the channel, floodplain, vegetation, flow or sediment David Penrose supply significantly affect this equilibrium, the stream may Todd St. John become unstable and start adjusting toward a new equilibrium state. -

Restoring Streams in an Urbanizing World
Freshwater Biology (2007) 52, 738–751 doi:10.1111/j.1365-2427.2006.01718.x Restoring streams in an urbanizing world EMILY S. BERNHARDT* AND MARGARET A. PALMER† *Department of Biology, Duke University, Durham, NC, U.S.A. †Chesapeake Biological Laboratory, University of Maryland Center for Environmental Science, Solomons, MD, U.S.A. SUMMARY 1. The world’s population is increasingly urban, and streams and rivers, as the low lying points of the landscape, are especially sensitive to and profoundly impacted by the changes associated with urbanization and suburbanization of catchments. 2. River restoration is an increasingly popular management strategy for improving the physical and ecological conditions of degraded urban streams. In urban catchments, management activities as diverse as stormwater management, bank stabilisation, channel reconfiguration and riparian replanting may be described as river restoration projects. 3. Restoration in urban streams is both more expensive and more difficult than restoration in less densely populated catchments. High property values and finely subdivided land and dense human infrastructure (e.g. roads, sewer lines) limit the spatial extent of urban river restoration options, while stormwaters and the associated sediment and pollutant loads may limit the potential for restoration projects to reverse degradation. 4. To be effective, urban stream restoration efforts must be integrated within broader catchment management strategies. A key scientific and management challenge is to establish criteria for determining when the design options for urban river restoration are so constrained that a return towards reference or pre-urbanization conditions is not realistic or feasible and when river restoration presents a viable and effective strategy for improving the ecological condition of these degraded ecosystems. -

1 Manitou Stream
LEAFLET 20 • May 2010 Manitou Stream Photo by Paul Zedler How Best to Manage “UrBan streaM syndroMe” Improving water quality: site with Arboretum staff on 23 April 2010. Accompanying her was another noted Newer, greener approaches stream ecologist, Dr. Bobbi Peckarsky, Across the nation, 37,099 river-restoration projects cost their Emeritus Professor from Cornell U., proponents and taxpayers an average of over $1 billion per now Honorary Fellow in Zoology at year (Bernhardt et al. 2005). Most had the goal of improving UW-Madison. water quality, but only a tiny minority had any monitoring How might Manitou Stream be data to allow comparison of benefits to costs—in either dollars rehabilitated? In North Carolina, or unintended impacts to the environment (ibid.). Among the Bernhardt’s data for engineered most degraded rivers and streams are those that flow through approaches to abating the urban stream urban areas. In Bernhardt’s terminology, urban streams are Dr. Emily Bernhardt solution indicate an average cost of over $1 “disconnected from their floodplains and hyperconnected to their million per project, with minimal benefit watersheds,” through drainage channels and stormwater pipes. (ecosystem services) when they fail to reconnect the stream to its The Arboretum’s Manitou Stream, near the intersection floodplain. Typical projects aim to remove obstructions to flow, of Nakoma Road and Manitou Way, fits this description. grade the banks (which releases sediment during construction), Following are answers to three questions: How would ecologists then stabilize streambeds and banks with riprap and turf restore Manitou Stream; how do engineers propose to control reinforcement matting. -

Quantitative Geomorphology of Drainage Basins Related to Fish
INFORMATIONAL LEAFLET NO. 162 QUANTITATIVE GEOMORPHOLOGY OF DRA INAGE BAS INS RELATED TO F ISH PRODUCT ION BY G. L. Ziemer STATE OF ALASKA William A. Egan - Governor DEPARTMENT OF F l SH AND GAME James W. Brooks, Commissioner Subport Building, Juneau 99801 July 1973 TABLE OF CONTENTS Page ABSTRACT .............................. 1 INTRODUCTION .......................... 1 GEOMORPHIC ELEMENTS ...................... 2 OBJECTIVES ............................ 4 METHODOLOGY .......................... 4 SUMMARY ............................. 11 CONCLUSIONS ........................... 14 REFERENCES ............................ 18 APPENDIX A . GEOMORPHIC ELEMENTS .............. 19 APPENDIX B . MEASURE OF FISH PRODUCTION .......... 25 QUANTITATIVE GEOMORPHOLOCX OF DRAINAGE BASINS RELATED TO FISH PRODUCTION G.L. Ziemer, P.E. Chief Engineer Alaska Department of Fish and Game Juneau, Alaska ABSTRACT This report covers the results of a study investigating the possibility of developing a classification index system for watersheds which would quan- tify their total composite salmon production potential. The premise was tested that, within a geologically and climatologically homogenous region, the water flow regimen of streams, and the channels that flow builds, is universally related to certain identifiable characteristics of their basins and drainages and that these control or indicate the level of fisheries production. This study shows that a correlation between drainage system geometry and freshwater production factors for anadromous fishes can be shown, and an index expressing that relationship, in the case of pink salmon in Prince William Sound, has been developed. INTRODUCTION As anadromous fisheries management proceeds from the basic position of husbandry of the existing stocks to the addition of programs designed to increase the quantity and to enhance the quality of the freshwater environment for fish production, it becomes desirable to provide to the manager better tools to equate one site against another so his projects return the maximum dividends. -

National Stream Internet Protocol and User Guide
National Stream Internet Protocol and User Guide David Nagel1, Erin Peterson2, Daniel Isaak1, Jay Ver Hoef3, and Dona Horan1 U.S. Forest Service, Rocky Mountain Research Station Air, Water, and Aquatic Environments Program 322 E. Front St., Boise, ID 1 Author affiliations: 1 US Forest Service, Rocky Mountain Research Station, AWAE Program, 322 E. Front St., Suite 401, Boise, ID 83702. 2 Queensland University of Technology, Brisbane, Queensland, Australia. 3 National Oceanic and Atmospheric Administration, Fairbanks, AK. Version 3-22-2017 Abstract The rate at which new information about stream resources is being created has accelerated with the recent development of spatial stream-network models (SSNMs), the growing availability of stream databases, and ongoing advances in geospatial science and computational efficiency. To further enhance information development, the National Stream Internet (NSI) project was developed as a means of providing a consistent, flexible analytical infrastructure that can be applied with many types of stream data anywhere in the country. A key part of that infrastructure is the NSI network, a digital GIS layer which has a specific topological structure that was designed to work effectively with SSNMs. The NSI network was derived from the National Hydrography Dataset Plus, Version 2 (NHDPlusV2) following technical procedures that ensure compatibility with SSNMs. This report describes those procedures and additional steps that are required to prepare datasets for use with SSNMs. 2 1.0 Introduction 1.1 Overview The USGS National Hydrography Dataset Plus, Version 2 (NHDPlusV2) (McKay et al., 2012) is an attribute rich, GIS stream network developed at 1:100,000 scale by the Environmental Protection Agency (EPA) and U.S. -

Spatial Patterns in CO 2 Evasion from the Global River Network Ronny Lauerwald, Goulven Laruelle, Jens Hartmann, Philippe Ciais, Pierre A.G
Spatial patterns in CO 2 evasion from the global river network Ronny Lauerwald, Goulven Laruelle, Jens Hartmann, Philippe Ciais, Pierre A.G. Regnier To cite this version: Ronny Lauerwald, Goulven Laruelle, Jens Hartmann, Philippe Ciais, Pierre A.G. Regnier. Spatial patterns in CO 2 evasion from the global river network. Global Biogeochemical Cycles, American Geophysical Union, 2015, 29 (5), pp.534 - 554. 10.1002/2014GB004941. hal-01806195 HAL Id: hal-01806195 https://hal.archives-ouvertes.fr/hal-01806195 Submitted on 28 Oct 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. PUBLICATIONS Global Biogeochemical Cycles RESEARCH ARTICLE Spatial patterns in CO2 evasion from the global 10.1002/2014GB004941 river network Key Points: Ronny Lauerwald1,2,3, Goulven G. Laruelle1,4, Jens Hartmann3, Philippe Ciais5, and Pierre A. G. Regnier1 • First global maps of river CO2 partial pressures and evasion at 0.5° 1Department of Earth and Environmental Sciences, Université Libre de Bruxelles, Brussels, Belgium, 2Institut Pierre-Simon resolution Laplace, Paris, France, 3Institute for Geology, University of Hamburg, Hamburg, Germany, 4Department of Earth Sciences- • Global river CO2 evasion estimated at À 5 650 (483–846) Tg C yr 1 Geochemistry, Utrecht University, Utrecht, Netherlands, LSCE IPSL, Gif Sur Yvette, France • Latitudes between 10°N and 10°S contribute half of the global CO2 evasion Abstract CO2 evasion from rivers (FCO2) is an important component of the global carbon budget. -

Fluvial Geomorphology
East Branch Delaware River Stream Corridor Management Plan Volume 2 Fluvial Geomorphology: the Hydrology: the science study of riverine landforms and dealing with the properties, the processes that create them distribution, and circulation of water on and below the earth’s surface and in the atmosphere (Merriam-Webster Online 09/12/07) An understanding of both hydrology and fluvial geomorphology is essential when approaching stream management at any level. This section is intended to serve as an overview of both aspects of stream science, as well as their relation to management practices past and present. Applied fluvial geomorphology utilizes the relationships and principles developed through the study of rivers and streams and how they function within their landscape to preserve and restore stream systems. In landscapes unchanged by human activities, streams reflect the regional climate, biology and geology. Bedrock and glacial deposits influence the stream system within its drainage basin. The “dendritic” (formed like the branches of a tree, Figure 3.1) stream pattern of the East Branch Delaware River watershed developed because horizontally bedded, sedimentary Figure 3.1 Stream Ordering (NRCS) and the bedrock had a gently sloping regional Depiction of a Dendritic Stream System dip at the time the initial drainage channels began forming16. The bedrock’s jointing pattern (the pattern of deep, vertical fractures) also influence stream pattern formation. The region’s geologic history has favored the development of non-symmetric drainage basins in the East Branch basin. As rivers flow across the landscape, they generally increase in size, merging with other rivers. This increase in size brings about a concept known as stream order. -
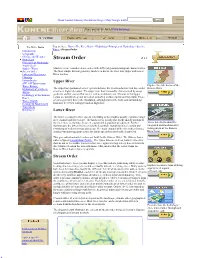
Stream Order
About Tutorial Glossary Documents Images Maps Google Earth Please provide feedback! Click for details The River Basin You are here: Home>The River Basin >Hydrology>Principles of Hydrology >Surface Water >Stream Order Introduction Geography Climate and Weather Hydrology Stream Order Principles of Hydrology Water Cycle Surface Water Most rivers are considered as reaches with different geomorphological characteristics. Stream Order The most simple divison generally made is to divide the river into Upper and Lower Lakes and Reservoirs River reaches Flooding Groundwater Upper River SW/ GW Interactions Water Balance Explore the sub- basins of the Hydrology of Southern The uppermost portion of a river system includes the river headwaters and low- order Kunene River Africa streams at higher elevation. The upper river basin is usually characterised by steep Hydrology of the Kunene gradients and by erosion that carries sediment downstream. Streams in this upper Basin region are usually steep and torrential, and often include rapids and waterfalls. These Water Quality streams generally have little floodplain, although part of the bank and surrounding Ecology & Biodiversity land may be wetted during periods of high flow. Watersheds References Lower River The lower section of a river system (extending to the mouth), usually exhibits a larger river channel and lower slope – the landscape is usually flat. In the middle portion of the river there is a balance between erosion and deposition of sediment. Further Video Interviews about the downstream, the lower river sees mainly deposition, though localised erosion and integrated and transboundary reworking of sediments may also occur. The main channel of the river often forms a management of the Kunene sinuous (meandering) path across the landscape unless artificially channelled. -

Synthetic Chemicals As Agents of Global Change
CONCEPTS AND QUESTIONS 84 Synthetic chemicals as agents of global change Emily S Bernhardt1*†, Emma J Rosi2†, and Mark O Gessner3,4 Though concerns about the proliferation of synthetic chemicals – including pesticides – gave rise to the modern environmental movement in the early 1960s, synthetic chemical pollution has not been included in most analyses of global change. We examined the rate of change in the production and variety of pesticides, pharmaceuticals, and other synthetic chemicals over the past four decades. We compared these rates to those for well- recognized drivers of global change such as rising atmospheric CO2 concentrations, nutrient pollu- tion, habitat destruction, and biodiversity loss. Our analysis showed that increases in synthetic chemical production and diversification, particularly within the developing world, outpaced these other agents of global change. Despite these trends, mainstream ecological journals, ecological meetings, and ecological funding through the US National Science Foundation devote less than 2% of their journal pages, meeting talks, and science funding, respectively, to the study of synthetic chemicals. Front Ecol Environ 2017; 15(2): 84–90, doi:10.1002/fee.1450 hen Rachel Carson wrote “The most alarming of all have been recognized as one of the critical markers of Wman’s assaults upon the environment is the what defines the modern era as the Anthropocene contamination of air, earth, water, and sea with dangerous (Waters et al. 2016), a new geologic epoch. Many of these and even lethal materials” (Carson 1962), she raised grave novel chemical entities are pesticides and pharmaceuti- concerns about the proliferation of pesticides in the US. cals – organic chemicals that are specifically designed to At that time there were 200 pesticides on the US market, kill or prevent the growth of unwanted organisms (weeds, and the World Health Organization estimated that nearly pathogens, pests), or to interfere with organismal bio- one million metric tons of pesticides were being applied to chemistry (UNEP 2013). -

Ecosystem Services in Practice: Management Decisions in the Public Sector— from Theory to Application
Seminar 6 Ecosystem Services in Practice: Management Decisions in the Public Sector— From Theory to Application Speaker Heather Wright · Dr. Carl Shapiro · Dr. Lydia Olander · Dr. Mary Ruckleshaus Ecosystem Services and Public Sector Decision-Making Introduction and Conclusion by Heather Wright, Gordon and Betty Moore Foundation INTRODUCTION Given the magnitude of the impact of global climate change and other human activities on our natural systems, there is a critical need for governments to support sustained and continued ecosystem benefits and to create incentives to maintain environmental capital. Traditionally, economic development goals have depended heavily on ecosystem services, but economic development activities tend to ignore the welfare of those ecosystems and thus jeopardize the well-being of people. This neglect of ecosystem services can increase ecological, social, and economic problems at local and global scales over the long term. As a result, long-term sustainability goals may be foregone for short-term economic wins. Linking policy and/or management objectives to ecosystem service objectives should and can be done in a way that aims to maximize economic, ecological, and social outcomes. To this end, decision-makers need to deliberately take into account the connections between development, ecosystems, and services provided. This particular seminar is devoted to looking more closely at how ecosystem services are considered in public sector decision-making. The examples showcased explore current ecosystem service approaches that are being applied in the public sector, and examine the rationale and incentives shaping management decisions. They also elucidate the key successes and shortcomings of their implementation. As demonstrated in the following cases, a pure command-and-control approach to mitigation is evolving into policies with a market-like mechanism (e.g., water funds, mitigation banks). -

A Mass Balance of Nitrogen in a Large Lowland River (Elbe, Germany)
water Article A Mass Balance of Nitrogen in a Large Lowland River (Elbe, Germany) , Stephanie Ritz * y and Helmut Fischer Federal Institute of Hydrology—BfG, Am Mainzer Tor 1, 56068 Koblenz, Germany; [email protected] * Correspondence: [email protected] Current address: Federal Agency of Nature Conservation—BfN, Konstantinstrasse 110, 53179 Bonn, Germany. y Received: 23 September 2019; Accepted: 13 November 2019; Published: 14 November 2019 Abstract: Nitrogen (N) delivered by rivers causes severe eutrophication in many coastal waters, and its turnover and retention are therefore of major interest. We set up a mass balance along a 582 km river section of a large, N-rich lowland river to quantify N retention along this river segment and to identify the underlying processes. Our assessments are based on four Lagrangian sampling campaigns performed between 2011 and 2013. Water quality data served as a basis for calculations of N retention, while chlorophyll-a and zooplankton counts were used to quantify the respective primary and secondary transformations of dissolved inorganic N into biomass. The mass balance 2 1 revealed an average N retention of 17 mg N m− h− for both nitrate N (NO3–N) and total N (TN). Stoichiometric estimates of the assimilative N uptake revealed that, although NO3–N retention was associated with high phytoplankton assimilation, only a maximum of 53% of NO3–N retention could be attributed to net algal assimilation. The high TN retention rates in turn were most probably caused by a combination of seston deposition and denitrification. The studied river segment acts as a TN sink by retaining almost 30% of the TN inputs, which shows that large rivers can contribute considerably to N retention during downstream transport.