The Skull O Neurocranium, Form and Function O Dermatocranium, Form
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
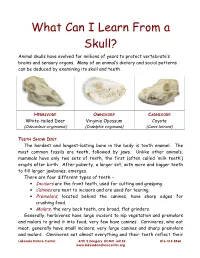
What Can I Learn from a Skull? Animal Skulls Have Evolved for Millions of Years to Protect Vertebrate’S Brains and Sensory Organs
What Can I Learn From a Skull? Animal skulls have evolved for millions of years to protect vertebrate’s brains and sensory organs. Many of an animal’s dietary and social patterns can be deduced by examining its skull and teeth. HERBIVORE OMNIVORE CARNIVORE White-tailed Deer Virginia Opossum Coyote (Odocoileus virginianus) (Didelphis virginiana) (Canis latrans) TEETH SHOW DIET . The hardest and longest-lasting bone in the body is tooth enamel. The most common fossils are teeth, followed by jaws. Unlike other animals, mammals have only two sets of teeth, the first (often called ‘milk teeth’) erupts after birth. After puberty, a larger set, with more and bigger teeth to fill larger jawbones, emerges. There are four different types of teeth – • Incisors are the front teeth, used for cutting and grasping. • Canines are next to incisors and are used for tearing. • Premolars , located behind the canines, have sharp edges for crushing food. • Molars , the very back teeth, are broad, flat grinders. Generally, herbivores have large incisors to nip vegetation and premolars and molars to grind it into food; very few have canines. Carnivores, who eat meat, generally have small incisors, very large canines and sharp premolars and molars. Omnivores eat almost everything and their teeth reflect their Lakeside Nature Center 4701 E Gregory, KCMO 64132 816-513-8960 www.lakesidenaturecenter.org preferences; they have all four types of teeth. In fact, if you’d like to see an excellent example of an omnivore’s teeth, look in the mirror. EYE PLACEMENT IDENTIFIES PREDATORS . Carnivores generally have large eyes, placed so that the eyes look forward and the areas of vision of the two eyes overlap. -
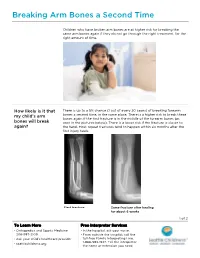
PE2812 Breaking Arm Bones a Second Time
Breaking Arm Bones a Second Time Children who have broken arm bones are at higher risk for breaking the same arm bones again if they do not go through the right treatment, for the right amount of time. How likely is it that There is up to a 5% chance (1 out of every 20 cases) of breaking forearm my child’s arm bones a second time, in the same place. There is a higher risk to break these bones again if the first fracture is in the middle of the forearm bones (as bones will break seen in the pictures below). There is a lower risk if the fracture is closer to again? the hand. Most repeat fractures tend to happen within six months after the first injury heals. First fracture Same fracture after healing for about 6 weeks 1 of 2 To Learn More Free Interpreter Services • Orthopedics and Sports Medicine • In the hospital, ask your nurse. 206-987-2109 • From outside the hospital, call the • Ask your child’s healthcare provider toll-free Family Interpreting Line, 1-866-583-1527. Tell the interpreter • seattlechildrens.org the name or extension you need. Breaking Arm Bones a Second Time How can I help my Wearing a cast for at least six weeks lowers the risk of breaking the same child lower the risk arm bones again. After wearing a cast, we recommend your child wear a brace for 4 weeks in order to protect the injured area and start improving of having a wrist movement. While your child wears a brace, we recommend they do repeated bone not participate in contact sports (e.g., soccer, football or dodge ball). -

Fishes of Terengganu East Coast of Malay Peninsula, Malaysia Ii Iii
i Fishes of Terengganu East coast of Malay Peninsula, Malaysia ii iii Edited by Mizuki Matsunuma, Hiroyuki Motomura, Keiichi Matsuura, Noor Azhar M. Shazili and Mohd Azmi Ambak Photographed by Masatoshi Meguro and Mizuki Matsunuma iv Copy Right © 2011 by the National Museum of Nature and Science, Universiti Malaysia Terengganu and Kagoshima University Museum All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means without prior written permission from the publisher. Copyrights of the specimen photographs are held by the Kagoshima Uni- versity Museum. For bibliographic purposes this book should be cited as follows: Matsunuma, M., H. Motomura, K. Matsuura, N. A. M. Shazili and M. A. Ambak (eds.). 2011 (Nov.). Fishes of Terengganu – east coast of Malay Peninsula, Malaysia. National Museum of Nature and Science, Universiti Malaysia Terengganu and Kagoshima University Museum, ix + 251 pages. ISBN 978-4-87803-036-9 Corresponding editor: Hiroyuki Motomura (e-mail: [email protected]) v Preface Tropical seas in Southeast Asian countries are well known for their rich fish diversity found in various environments such as beautiful coral reefs, mud flats, sandy beaches, mangroves, and estuaries around river mouths. The South China Sea is a major water body containing a large and diverse fish fauna. However, many areas of the South China Sea, particularly in Malaysia and Vietnam, have been poorly studied in terms of fish taxonomy and diversity. Local fish scientists and students have frequently faced difficulty when try- ing to identify fishes in their home countries. During the International Training Program of the Japan Society for Promotion of Science (ITP of JSPS), two graduate students of Kagoshima University, Mr. -

Study Guide Medical Terminology by Thea Liza Batan About the Author
Study Guide Medical Terminology By Thea Liza Batan About the Author Thea Liza Batan earned a Master of Science in Nursing Administration in 2007 from Xavier University in Cincinnati, Ohio. She has worked as a staff nurse, nurse instructor, and level department head. She currently works as a simulation coordinator and a free- lance writer specializing in nursing and healthcare. All terms mentioned in this text that are known to be trademarks or service marks have been appropriately capitalized. Use of a term in this text shouldn’t be regarded as affecting the validity of any trademark or service mark. Copyright © 2017 by Penn Foster, Inc. All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the copyright owner. Requests for permission to make copies of any part of the work should be mailed to Copyright Permissions, Penn Foster, 925 Oak Street, Scranton, Pennsylvania 18515. Printed in the United States of America CONTENTS INSTRUCTIONS 1 READING ASSIGNMENTS 3 LESSON 1: THE FUNDAMENTALS OF MEDICAL TERMINOLOGY 5 LESSON 2: DIAGNOSIS, INTERVENTION, AND HUMAN BODY TERMS 28 LESSON 3: MUSCULOSKELETAL, CIRCULATORY, AND RESPIRATORY SYSTEM TERMS 44 LESSON 4: DIGESTIVE, URINARY, AND REPRODUCTIVE SYSTEM TERMS 69 LESSON 5: INTEGUMENTARY, NERVOUS, AND ENDOCRINE S YSTEM TERMS 96 SELF-CHECK ANSWERS 134 © PENN FOSTER, INC. 2017 MEDICAL TERMINOLOGY PAGE III Contents INSTRUCTIONS INTRODUCTION Welcome to your course on medical terminology. You’re taking this course because you’re most likely interested in pursuing a health and science career, which entails proficiencyincommunicatingwithhealthcareprofessionalssuchasphysicians,nurses, or dentists. -

Development of the Muscles Associated with the Mandibular and Hyoid Arches in the Siberian Sturgeon, Acipenser Baerii (Acipenseriformes: Acipenseridae)
Received: 31 May 2017 | Revised: 24 September 2017 | Accepted: 29 September 2017 DOI: 10.1002/jmor.20761 RESEARCH ARTICLE Development of the muscles associated with the mandibular and hyoid arches in the Siberian sturgeon, Acipenser baerii (Acipenseriformes: Acipenseridae) Peter Warth1 | Eric J. Hilton2 | Benjamin Naumann1 | Lennart Olsson1 | Peter Konstantinidis3 1Institut fur€ Spezielle Zoologie und Evolutionsbiologie mit Phyletischem Abstract Museum, Friedrich-Schiller-Universität Jena, The skeleton of the jaws and neurocranium of sturgeons (Acipenseridae) are connected only Germany through the hyoid arch. This arrangement allows considerable protrusion and retraction of the 2 Department of Fisheries Science, Virginia jaws and is highly specialized among ray-finned fishes (Actinopterygii). To better understand the Institute of Marine Science, College of unique morphology and the evolution of the jaw apparatus in Acipenseridae, we investigated the William & Mary, Gloucester Point, Virginia development of the muscles of the mandibular and hyoid arches of the Siberian sturgeon, Aci- 3Department of Fisheries and Wildlife, Oregon State University, Corvallis, Oregon penser baerii. We used a combination of antibody staining and formalin-induced fluorescence of tissues imaged with confocal microscopy and subsequent three-dimensional reconstruction. These Correspondence data were analyzed to address the identity of previously controversial and newly discovered mus- Peter Warth, Institut fur€ Spezielle Zoologie cle portions. Our results indicate that the anlagen of the muscles in A. baerii develop similarly to und Evolutionsbiologie mit Phyletischem Museum, Friedrich-Schiller-Universität Jena, those of other actinopterygians, although they differ by not differentiating into distinct muscles. Erbertstr. 1, 07743 Jena, Germany. This is exemplified by the subpartitioning of the m. adductor mandibulae as well as the massive m. -

The Ear in Mammal-Like Reptiles and Early Mammals
Acta Palaeontologica Polonica Vol. 28, No. 1-2 pp, 147-158 Warszawa, 1983 Second Symposium on Mesozoic T erre stial Ecosystems, Jadwisin 1981 KENNETH A. KERMACK and FRANCES MUSSETT THE EAR IN MAMMAL-LIKE REPTILES AND EARLY MAMMALS KERMACK, K . A. a nd MUSS ETT, F.: The ear in mammal-like r eptiles an d early mammals. Acta Palaeont. P olonica , 28, 1-2, 147-158, 1983. Th e early m embers of the Theropsida lacked a tympanic membrane. In the later theropslds, the Therapsid a, a tym p an ic membrane develop ed from thc skin on the lateral side of th e lower jaw. The tympanum is not homologous In the Therapsida and ' t he Sauropslda. The ther apsid ea r w as a poor receiver of airborne sound, both In hi gh frequency r esp onse and In the r ange of frequencies encompassed. With the radiation of the Sauropsida in the Triassic the large therapsids became extinct, the small therap si ds evolv ed In to the mammal s and became nocturnal. High frequency hearin g w as essen tial for the nocturn al mode of life; quadrate and arttcutar became diss ociated from the jaw hinge to become the m ammali an au di tory ossi cles . I n the Theria the cochlea became coil ed. The spiral cochlea could n ot have existed until there w as a middle ear w ith the n ec essary h ig h f re q uency r esp onse. This m ay n ot have been until the Cretace ous. -

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V. -

GLOSSARY of MEDICAL and ANATOMICAL TERMS
GLOSSARY of MEDICAL and ANATOMICAL TERMS Abbreviations: • A. Arabic • abb. = abbreviation • c. circa = about • F. French • adj. adjective • G. Greek • Ge. German • cf. compare • L. Latin • dim. = diminutive • OF. Old French • ( ) plural form in brackets A-band abb. of anisotropic band G. anisos = unequal + tropos = turning; meaning having not equal properties in every direction; transverse bands in living skeletal muscle which rotate the plane of polarised light, cf. I-band. Abbé, Ernst. 1840-1905. German physicist; mathematical analysis of optics as a basis for constructing better microscopes; devised oil immersion lens; Abbé condenser. absorption L. absorbere = to suck up. acervulus L. = sand, gritty; brain sand (cf. psammoma body). acetylcholine an ester of choline found in many tissue, synapses & neuromuscular junctions, where it is a neural transmitter. acetylcholinesterase enzyme at motor end-plate responsible for rapid destruction of acetylcholine, a neurotransmitter. acidophilic adj. L. acidus = sour + G. philein = to love; affinity for an acidic dye, such as eosin staining cytoplasmic proteins. acinus (-i) L. = a juicy berry, a grape; applied to small, rounded terminal secretory units of compound exocrine glands that have a small lumen (adj. acinar). acrosome G. akron = extremity + soma = body; head of spermatozoon. actin polymer protein filament found in the intracellular cytoskeleton, particularly in the thin (I-) bands of striated muscle. adenohypophysis G. ade = an acorn + hypophyses = an undergrowth; anterior lobe of hypophysis (cf. pituitary). adenoid G. " + -oeides = in form of; in the form of a gland, glandular; the pharyngeal tonsil. adipocyte L. adeps = fat (of an animal) + G. kytos = a container; cells responsible for storage and metabolism of lipids, found in white fat and brown fat. -

6. the Pharynx the Pharynx, Which Forms the Upper Part of the Digestive Tract, Consists of Three Parts: the Nasopharynx, the Oropharynx and the Laryngopharynx
6. The Pharynx The pharynx, which forms the upper part of the digestive tract, consists of three parts: the nasopharynx, the oropharynx and the laryngopharynx. The principle object of this dissection is to observe the pharyngeal constrictors that form the back wall of the vocal tract. Because the cadaver is lying face down, we will consider these muscles from the back. Figure 6.1 shows their location. stylopharyngeus suuperior phayngeal constrictor mandible medial hyoid bone phayngeal constrictor inferior phayngeal constrictor Figure 6.1. Posterior view of the muscles of the pharynx. Each of the three pharyngeal constrictors has a left and right part that interdigitate (join in fingerlike branches) in the midline, forming a raphe, or union. This raphe forms the back wall of the pharynx. The superior pharyngeal constrictor is largely in the nasopharynx. It has several origins (some texts regard it as more than one muscle) one of which is the medial pterygoid plate. It assists in the constriction of the nasopharynx, but has little role in speech production other than helping form a site against which the velum may be pulled when forming a velic closure. The medial pharyngeal constrictor, which originates on the greater horn of the hyoid bone, also has little function in speech. To some extent it can be considered as an elevator of the hyoid bone, but its most important role for speech is simply as the back wall of the vocal tract. The inferior pharyngeal constrictor also performs this function, but plays a more important role constricting the pharynx in the formation of pharyngeal consonants. -

98796-Anatomy of the Orbit
Anatomy of the orbit Prof. Pia C Sundgren MD, PhD Department of Diagnostic Radiology, Clinical Sciences, Lund University, Sweden Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 Lay-out • brief overview of the basic anatomy of the orbit and its structures • the orbit is a complicated structure due to its embryological composition • high number of entities, and diseases due to its composition of ectoderm, surface ectoderm and mesoderm Recommend you to read for more details Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 3 x 3 Imaging technique 3 layers: - neuroectoderm (retina, iris, optic nerve) - surface ectoderm (lens) • CT and / or MR - mesoderm (vascular structures, sclera, choroid) •IOM plane 3 spaces: - pre-septal •thin slices extraconal - post-septal • axial and coronal projections intraconal • CT: soft tissue and bone windows 3 motor nerves: - occulomotor (III) • MR: T1 pre and post, T2, STIR, fat suppression, DWI (?) - trochlear (IV) - abducens (VI) Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 Superior orbital fissure • cranial nerves (CN) III, IV, and VI • lacrimal nerve • frontal nerve • nasociliary nerve • orbital branch of middle meningeal artery • recurrent branch of lacrimal artery • superior orbital vein • superior ophthalmic vein Lund University / Faculty of Medicine / Inst. Clinical Sciences / Radiology / ECNR Dubrovnik / Oct 2018 Lund University / Faculty of Medicine / Inst. -

Morphology of the Foramen Magnum in Young Eastern European Adults
Folia Morphol. Vol. 71, No. 4, pp. 205–216 Copyright © 2012 Via Medica O R I G I N A L A R T I C L E ISSN 0015–5659 www.fm.viamedica.pl Morphology of the foramen magnum in young Eastern European adults F. Burdan1, 2, J. Szumiło3, J. Walocha4, L. Klepacz5, B. Madej1, W. Dworzański1, R. Klepacz3, A. Dworzańska1, E. Czekajska-Chehab6, A. Drop6 1Department of Human Anatomy, Medical University of Lublin, Lublin, Poland 2St. John’s Cancer Centre, Lublin, Poland 3Department of Clinical Pathomorphology, Medical University of Lublin, Lublin, Poland 4Department of Anatomy, Collegium Medicum, Jagiellonian University, Krakow, Poland 5Department of Psychiatry and Behavioural Sciences, Behavioural Health Centre, New York Medical College, Valhalla NY, USA 6Department of General Radiology and Nuclear Medicine, Medical University of Lublin, Lublin, Poland [Received 21 July 2012; Accepted 7 September 2012] Background: The foramen magnum is an important anatomical opening in the base of the skull through which the posterior cranial fossa communicates with the vertebral canal. It is also related to a number of pathological condi- tions including Chiari malformations, various tumours, and occipital dysplasias. The aim of the study was to evaluate the morphology of the foramen magnum in adult individuals in relation to sex. Material and methods: The morphology of the foramen magnum was evalu- ated using 3D computer tomography images in 313 individuals (142 male, 171 female) aged 20–30 years. Results: The mean values of the foramen length (37.06 ± 3.07 vs. 35.47 ± ± 2.60 mm), breadth (32.98 ± 2.78 vs. 30.95 ± 2.71 mm) and area (877.40 ± ± 131.64 vs. -

MORPHOMETRIC and MORPHOLOGICAL ANALYSIS of FORAMEN OVALE in DRY HUMAN SKULLS Ashwini
International Journal of Anatomy and Research, Int J Anat Res 2017, Vol 5(1):3547-51. ISSN 2321-4287 Original Research Article DOI: https://dx.doi.org/10.16965/ijar.2017.109 MORPHOMETRIC AND MORPHOLOGICAL ANALYSIS OF FORAMEN OVALE IN DRY HUMAN SKULLS Ashwini. N.S *1, Venkateshu. K.V 2. *1 Assistant Professor, Department Of Anatomy,Sri Devaraj Urs Medical College ,Tamaka, Kolar, Karnataka, India. 2 Professor And Head, Department Of Anatomy ,Sri Devaraj Urs Medical College, Tamaka, Kolar, India. ABSTRACT Introduction: Foramen ovale is an important foramen in the middle cranial fossa. Foramen ovale is situated in the greater wing of sphenoid bone, posterior to the foramen rotandum. Through the foramen ovale the mandibular nerve, accessory meningeal artery, lesser petrosal nerve, emissary veins pass .The shape of the foramen ovale is usually oval compared to other foramen of the skull. Materials and Methods: The study was conducted on 55 dry human skulls(110 sides) in Department of Anatomy, Sri Devaraj Urs Medical college, Tamaka, Kolar. Skulls in poor condition or skulls with partially damaged surroundings around the foramen ovale were excluded from the study. Linear measurements were taken on right and left sides of foramen ovale using divider and meter rule. Results: The maximum length of foramen ovale was 14 mm on the right side and 17 mm on the left, its maximum breadth on the right side was 8mm and 10mm on the left . Through the statistical analysis of morphometric measurements between right and left foramen ovale which was found to be insignificant , the results of both sides marks as the evidence of asymmetry in the morphometry of the foramen ovale.