On the Nature of Simultaneous Colour Contrast
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

When Red Lights Look Yellow
Publications 11-2005 When Red Lights Look Yellow Joanne M. Wood Queensland University of Technology David A. Atchison Queensland University of Technology Alex Chaparro Wichita State University, [email protected] Follow this and additional works at: https://commons.erau.edu/publication Part of the Cognition and Perception Commons, Musculoskeletal, Neural, and Ocular Physiology Commons, and the Ophthalmology Commons Scholarly Commons Citation Wood, J. M., Atchison, D. A., & Chaparro, A. (2005). When Red Lights Look Yellow. Investigative Ophthalmology & Visual Science, 46(11). https://doi.org/10.1167/iovs.04-1513 This Article is brought to you for free and open access by Scholarly Commons. It has been accepted for inclusion in Publications by an authorized administrator of Scholarly Commons. For more information, please contact [email protected]. When Red Lights Look Yellow Joanne M. Wood,1 David A. Atchison,1 and Alex Chaparro2 PURPOSE. Red signals are typically used to signify danger. This observer’s distance spectacle prescription. It did not occur study was conducted to investigate a situation identified by with lens powers in excess of ϩ1.00 D, as the signals became train drivers in which red signals appear yellow when viewed too blurred for the viewer to distinguish the color. A compre- at long distances (ϳ900 m) through progressive-addition hensive eye and vision examination of the train driver who had lenses. originally reported the color misperception revealed that his METHODS. A laboratory study was conducted to investigate the corrected vision was normal. The train driver was also shown effects of defocus, target size, ambient illumination, and sur- to have normal color vision, as assessed by the Ishihara, Farns- round characteristics on the extent of the color misperception worth Lantern, and Farnsworth D15 tests. -

Several Color Appearance Phenomena in Color Reproduction
2nd International Conference on Electronic & Mechanical Engineering and Information Technology (EMEIT-2012) Several Color Appearance Phenomena in Color Reproduction Qin-ling Dai1,a, Xiao-zhou Li2*,b, Ai Xu2 1 School of Materials Engineering (Southwest Forestry University), Kunming, China 650224 2 Key Laboratory of Pulp & Paper Science and Technology (Shandong Polytechnic University), Ministry of Education, Ji’nan, China, 250353 [email protected], bcorresponding author: [email protected] Keywords: color reproduction, color appearance, color appearance phenomena Abstract. Color perceived performance was influenced by various color appearance phenomena caused by varying viewing conditions in color reproduction process. It is necessary to do some research on the color appearance phenomena to represent the color appearance models qualitatively and quantitatively and accurate color reproduction easily. Only the phenomena were studied thoroughly, could the color transmission and reproduction be well performed. The color appearance and common color appearance phenomena of color reproduction were analyzed in this paper. And the basic theory of color appearance in color reproduction was also studied. Introduction High fidelity digital printing plays an important role in high fidelity color transmission and reproduction and it is one of the most important techniques to perform high fidelity color reproduction. High fidelity digital printing helps to perform accurate color reproduction of the original which can’t be performed because of paper and ink in traditional printing process [1]. In color printing, the color difference caused by paper, ink and viewing conditions is various. The difference is not only colorimetric difference but also different color appearance phenomena. While the different color appearance phenomenon is the leading factor to influence the color vision perceived. -

Auditory Experience Controls the Maturation of Song Discrimination and Sexual Response in Drosophila Xiaodong Li, Hiroshi Ishimoto, Azusa Kamikouchi*
RESEARCH ARTICLE Auditory experience controls the maturation of song discrimination and sexual response in Drosophila Xiaodong Li, Hiroshi Ishimoto, Azusa Kamikouchi* Graduate School of Science, Nagoya University, Nagoya, Japan Abstract In birds and higher mammals, auditory experience during development is critical to discriminate sound patterns in adulthood. However, the neural and molecular nature of this acquired ability remains elusive. In fruit flies, acoustic perception has been thought to be innate. Here we report, surprisingly, that auditory experience of a species-specific courtship song in developing Drosophila shapes adult song perception and resultant sexual behavior. Preferences in the song-response behaviors of both males and females were tuned by social acoustic exposure during development. We examined the molecular and cellular determinants of this social acoustic learning and found that GABA signaling acting on the GABAA receptor Rdl in the pC1 neurons, the integration node for courtship stimuli, regulated auditory tuning and sexual behavior. These findings demonstrate that maturation of auditory perception in flies is unexpectedly plastic and is acquired socially, providing a model to investigate how song learning regulates mating preference in insects. DOI: https://doi.org/10.7554/eLife.34348.001 Introduction Vocal learning in infants or juvenile birds relies heavily on the early experience of the adult conspe- cific sounds. In humans, early language input is necessary to form the ability of phonetic distinction *For correspondence: and pattern detection in the phase of auditory learning (Doupe and Kuhl, 1999; Kuhl, 2004). [email protected] Because of the strong parallels between speech acquisition of humans and song learning of song- Competing interests: The birds, and the difficulties to investigate the neural mechanisms of human early auditory memory at authors declare that no cellular resolution, songbirds have been used as a predominant model in studying memory formation competing interests exist. -
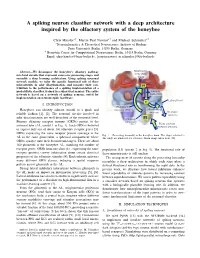
A Spiking Neuron Classifier Network with a Deep Architecture Inspired By
A spiking neuron classifier network with a deep architecture inspired by the olfactory system of the honeybee Chris Hausler¨ ∗†, Martin Paul Nawrot∗† and Michael Schmuker∗† ∗Neuroinformatics & Theoretical Neuroscience, Institute of Biology Freie Universitat¨ Berlin, 14195 Berlin, Germany † Bernstein Center for Computational Neuroscience Berlin, 10115 Berlin, Germany Email: [email protected], fmartin.nawrot, [email protected] Abstract—We decompose the honeybee’s olfactory pathway Mushroom into local circuits that represent successive processing stages and body (MB) calyx resemble a deep learning architecture. Using spiking neuronal network models, we infer the specific functional role of these microcircuits in odor discrimination, and measure their con- 2 tribution to the performance of a spiking implementation of a probabilistic classifier, trained in a supervised manner. The entire Kenyon network is based on a network of spiking neurons, suited for Cells (KC) implementation on neuromorphic hardware. Lateral horn 3 I. INTRODUCTION Projection Honeybees can identify odorant stimuli in a quick and neurons (PN) MB-extrinsic neurons To motor reliable fashion [1], [2]. The neuronal circuits involved in neurons odor discrimination are well described at the structural level. 1 Antennal Primary olfactory receptor neurons (ORNs) project to the lobe (AL) From odorant antennal lobe (AL, circuit 1 in Fig. 1). Each ORN is believed receptor neurons to express only one of about 160 olfactory receptor genes [3]. ORNs expressing the same receptor protein converge in the Fig. 1. Processing hierarchy in the honeybee brain. The stages relevant to AL in the same glomerulus, a spherical compartment where this study are numbered for reference. -

Role of Stereopsis Losses in Older Adults’ Performance
ROLE OF STEREOPSIS LOSSES IN OLDER ADULTS’ PERFORMANCE ON THE FINE-GRAIN MOVEMENT ILLUSION TASK by Marlena Pearson Honours Bachelor of Science, Psychology, Nipissing University, North Bay, Ontario, 2014 Honours Bachelor of Arts, Criminal Justice, Nipissing University, North Bay, Ontario, 2014 A thesis presented to Ryerson University in partial fulfillment of the requirement for the degree of Master of Arts in the program of Psychology Toronto, Ontario, Canada, 2017 © Marlena Pearson, 2017 Author’s Declaration AUTHOR'S DECLARATION FOR ELECTRONIC SUBMISSION OF A THESIS I hereby declare that I am the sole author of this thesis. This is a true copy of the thesis, including any required final revisions, as accepted by my examiners. I authorize Ryerson University to lend this thesis to other institutions or individuals for the purpose of scholarly research I further authorize Ryerson University to reproduce this thesis by photocopying or by other means, in total or in part, at the request of other institutions or individuals for the purpose of scholarly research. I understand that my thesis may be made electronically available to the public. ii Role of Stereopsis Losses in Older Adults’ Performance on the Fine-Grain Movement Illusion Task Master of Arts, 2017 Marlena Pearson Psychology, Ryerson University Abstract Accurate motion perception is necessary for older adults to safely navigate their environments. Yet it is not clear how stereopsis losses contribute to findings of motion perception deficits in older adults. To assess the contribution of stereopsis losses, three groups (younger adults, older adults with intact stereopsis, older adults with poor stereopsis) were recruited for a fine-grain movement task. -

1P MON. PM Cortex and Subcortical Auditory Nuclei
dynamic role for the vestibular system in orientation and flight control. Laboratory studies of flight behavior under illuminated and dark conditions in both static and rotating obstacle tests were carried out while administering heavy water ͑D2O͒ to bats to impair their vestibular inputs. Eptesicus carried out complex maneuvers through both fixed arrays of wires and a rotating obstacle array using both vision and echolocation, or when guided by echolocation alone. When treated with D2O in combination with lack of visual cues, bats showed considerable decrements in performance. These data indicate that big brown bats use both vision and echolocation to provide spatial registration for head position information generated by the vestibular system. 2:55 1pAB3. Bat’s auditory system: Corticofugal feedback and plasticity. Nobuo Suga ͑Dept. of Biol., Washington Univ., One Brookings Dr., St. Louis, MO 63130͒ The auditory system of the mustached bat consists of physiologically distinct subdivisions for processing different types of biosonar information. It was found that the corticofugal ͑descending͒ auditory system plays an important role in improving and adjusting auditory signal processing. Repetitive acoustic stimulation, cortical electrical stimulation or auditory fear conditioning evokes plastic changes of the central auditory system. The changes are based upon egocentric selection evoked by focused positive feedback associated with lateral inhibition. Focal electric stimulation of the auditory cortex evokes short-term changes in the auditory 1p MON. PM cortex and subcortical auditory nuclei. An increase in a cortical acetylcholine level during the electric stimulation changes the cortical changes from short-term to long-term. There are two types of plastic changes ͑reorganizations͒: centripetal best frequency shifts for expanded reorganization of a neural frequency map and centrifugal best frequency shifts for compressed reorganization of the map. -

Hue and Saturation Shifts from Spatially Induced Blackness
Bimler et al. Vol. 26, No. 1/January 2009/J. Opt. Soc. Am. A 163 Hue and saturation shifts from spatially induced blackness David L. Bimler,1,* Galina V. Paramei,2 and Chingis A. Izmailov3 1Department of Health and Human Development, Massey University, Private Bag 11-222, Palmerston North, New Zealand 2Department of Psychology, Liverpool Hope University, Hope Park, L16 9JD Liverpool, United Kingdom 3Department of Psychophysiology, Moscow Lomonosov State University, Mokhovaya st. 11/5, 125009 Moscow, Russia *Corresponding author: [email protected] Received June 24, 2008; revised September 15, 2008; accepted September 21, 2008; posted November 3, 2008 (Doc. ID 97849); published December 24, 2008 We studied changes in the color appearance of a chromatic stimulus as it underwent simultaneous contrast with a more luminous surround. Three normal trichromats provided color-naming descriptions for a 10 cd/m2 monochromatic field while a broadband white annulus surround ranged in luminance from 0.2 cd/m2 to 200 cd/m2. Descriptions of the chromatic field included Red, Green, Blue, Yellow, White, and Black or their combinations. The naming frequencies for each color/surround were used to calculate measures of similarity among the stimuli. Multidimensional scaling analysis of these subjective similarities resulted in a four-dimensional color space with two chromatic axes, red–green and blue–yellow, and two achromatic axes, revealing separate qualities of blackness/lightness and saturation. Contrast-induced darkening of the chro- matic field was found to be accompanied by shifts in both hue and saturation. Hue shifts were similar to the Bezold–Brücke shift; shifts in saturation were also quantified. -

Polarity Selectivity of Spatial Interactions in Perceived Contrast
Journal of Vision (2012) 12(2):3, 1–10 http://www.journalofvision.org/content/12/2/3 1 Polarity selectivity of spatial interactions in perceived contrast Department of Psychology, The University of Tokyo, Japan,& Human and Information Science Laboratory, Hiromi Sato NTT Communication Science Laboratories, NTT, Japan Human and Information Science Laboratory, Isamu Motoyoshi NTT Communication Science Laboratories, NTT, Japan Department of Psychology, Takao Sato The University of Tokyo, Japan The apparent contrast of a texture is reduced when surrounded by another texture with high contrast. This contrast–contrast phenomenon has been thought to be a result of spatial interactions between visual channels that encode contrast energy. In the present study, we show that contrast–contrast is selective to luminance polarity by using texture patterns composed of sparse elongated blobs. The apparent contrast of a texture of bright (dark) elements was substantially reduced only when it was surrounded by a texture of elements with the same polarity. This polarity specificity was not evident for textures with high element densities, which were similar to those used in previous studies, probably because such stimuli should inevitably activate both on- and off-type sensors. We also found that polarity-selective suppression decreased as the difference in orientation between the center and surround elements increased but remained for orthogonally oriented elements. These results suggest that the contrast–contrast illusion largely depends on spatial interactions between visual channels that are selective to on–off polarity and only weakly selective to orientation. Keywords: polarity selectivity, spatial interactions, perceived contrast Citation: Sato, H., Motoyoshi, I., & Sato, T. -

Visual Context Processing in Schizophrenia
Empirical Article Clinical Psychological Science 1(1) 5 –15 Visual Context Processing in Schizophrenia © The Author(s) 2013 Reprints and permission: sagepub.com/journalsPermissions.nav DOI: 10.1177/2167702612464618 http://cpx.sagepub.com Eunice Yang1,2, Duje Tadin3,4, Davis M. Glasser3, Sang Wook Hong1,5, Randolph Blake1,2, and Sohee Park1 1Department of Psychology, Vanderbilt University; 2Department of Brain and Cognitive Sciences, Seoul National University, Republic of Korea; 3Center for Visual Science and Department of Brain and Cognitive Sciences, University of Rochester; 4Department of Ophthalmology, University of Rochester; and 5Department of Psychology, Florida Atlantic University Abstract Abnormal perceptual experiences are central to schizophrenia, but the nature of these anomalies remains undetermined. We investigated contextual processing abnormalities across a comprehensive set of visual tasks. For perception of luminance, size, contrast, orientation, and motion, we quantified the degree to which the surrounding visual context altered a center stimulus’s appearance. Healthy participants showed robust contextual effects across all tasks, as evidenced by pronounced misperceptions of center stimuli. Schizophrenia patients exhibited intact contextual modulations of luminance and size but showed weakened contextual modulations of contrast, performing more accurately than controls. Strong motion and orientation context effects correlated with worse symptoms and social functioning. Importantly, the overall strength of contextual -

Krogh's Principle
Introduction to Neuroscience: Behavioral Neuroscience Neuroethology, Comparative Neuroscience, Natural Neuroscience Nachum Ulanovsky Department of Neurobiology, Weizmann Institute of Science 2017-2018, 2nd semester Principles of Neuroethology Neuroethology seeks to understand the mechanisms by which the Neurobiology central nervous system controls the Neuroethology Ethology natural behavior of animals. • Focus on Natural behaviors: Choosing to study a well-defined and reproducible yet natural behavior (either Innate or Learned behavior) • Need to study thoroughly the animal’s behavior, including in the field: Neuroethology starts with a good understanding of Ethology. • If you study the animals in the lab, you need to keep them in conditions as natural as possible, to avoid the occurrence of unnatural behaviors. • Krogh’s principle 1 Krogh’s principle August Krogh Nobel prize 1920 “For such a large number of problems there will be some animal of choice or a few such animals on which it can be most conveniently studied. Many years ago when my teacher, Christian Bohr, was interested in the respiratory mechanism of the lung and devised the method of studying the exchange through each lung separately, he found that a certain kind of tortoise possessed a trachea dividing into the main bronchi high up in the neck, and we used to say as a laboratory joke that this animal had been created expressly for the purposes of respiration physiology. I have no doubt that there is quite a number of animals which are similarly "created" for special physiological -

Lateral Inhibition Effects Demonstrate That the Maturational State of Neurons That Then Inhibit Their Neighbors
522 Lateral Inhibition effects demonstrate that the maturational state of neurons that then inhibit their neighbors. When a the learner’s brain is crucial for the attainment of stimulus (such as a bar of light or any other stim- a language system. ulus) excites a number of neurons in the network Several questions remain open about the mecha- (in this case neurons 4e, 5e, 6e, and 7e), the effect nisms underlying language learning. One issue is of inhibition is to suppress the neurons just out- whether specific aspects of language acquisition side the edge of the bar (3e and 8e) because those should be attributed to language-specific versus neurons are inhibited but not excited. Further, general-purpose learning mechanisms. Another issue because the neurons just inside the edges of the is whether children’s native language can affect the bar (4e and 7e) are excited by light and only inhib- way they think, and whether language is necessary ited by one neighbor, they are especially active. or helpful for the development of human concepts. This leads to perceptual contrast enhancement at borders. Further research showed that lateral inhi- Anna Papafragou bition also applied to overlapping stimuli, and that its strength fell off with distance between the See also Aphasias; Audition: Cognitive Influences; Context Effects in Perception; Speech Perception; Top- interacting stimuli. Down and Bottom-Up Processing; Word Recognition Haldan Keffer Hartline won the Nobel Prize in 1967 for discovering lateral inhibition and its neu- ral correlates. The first inhibitory circuit in the Further Readings nervous system was found in the horseshoe crab (Limulus polyphemus). -

Color Vision Mechanisms
11 COLOR VISION MECHANISMS Andrew Stockman Department of Visual Neuroscience UCL Institute of Opthalmology London, United KIngdom David H. Brainard Department of Psychology University of Pennsylvania Philadelphia, Pennsylvania 11.1 GLOSSARY Achromatic mechanism. Hypothetical psychophysical mechanisms, sometimes equated with the luminance mechanism, which respond primarily to changes in intensity. Note that achromatic mech- anisms may have spectrally opponent inputs, in addition to their primary nonopponent inputs. Bezold-Brücke hue shift. The shift in the hue of a stimulus toward either the yellow or blue invariant hues with increasing intensity. Bipolar mechanism. A mechanism, the response of which has two mutually exclusive types of out- put that depend on the balance between its two opposing inputs. Its response is nulled when its two inputs are balanced. Brightness. A perceptual measure of the apparent intensity of lights. Distinct from luminance in the sense that lights that appear equally bright are not necessarily of equal luminance. Cardinal directions. Stimulus directions in a three-dimensional color space that silence two of the three “cardinal mechanisms.” These are the isolating directions for the L+M, L–M, and S–(L+M) mech- anisms. Note that the isolating directions do not necessarily correspond to mechanism directions. Cardinal mechanisms. The second-site bipolar L–M and S–(L+M) chromatic mechanisms and the L+M luminance mechanism. Chromatic discrimination. Discrimination of a chromatic target from another target or back- ground, typically measured at equiluminance. Chromatic mechanism. Hypothetical psychophysical mechanisms that respond to chromatic stimuli, that is, to stimuli modulated at equiluminance. Color appearance. Subjective appearance of the hue, brightness, and saturation of objects or lights.