Comparison of Morphological and Anatomical Properties of Endangered Endemic Iris Pamphylica and I
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Šumarski Fakultet Preddiplomski Studij
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by University of Zagreb Repository ŠUMARSKI FAKULTET PREDDIPLOMSKI STUDIJ ŠUMARSTVO JOSIP ŽAČEK TAKSONOMSKI STATUS, RASPROSTRANJENOST I MORFOLOŠKE ZNAČAJKE AUTOHTONIH VRSTA RODA IRIS L. U HRVATSKOJ ZAVRŠNI RAD ZAGREB, rujan 2015. Zavod: Zavod za šumarsku genetiku, dendrologiju i botaniku Predmet: Šumarska botanika Mentor: Prof. dr. sc. Jozo Franjić Asistent: Dr. sc. Daniel Krstonošić Student: Josip Žaček JMBAG: 0068210787 Akad. godina: 2014/2015 Mjesto,datum Zagreb, 25. rujna 2015. godine obrane: Sadržaj rada: Slika: 18 Tablica: 0 Navoda literature: 10 Sažetak: Rod Iris L. je rasprostranjen na Sjevernoj polutci. Rod se odlikuje velikim brojem vrsta, podvrsta, varijeteta i križanaca. Za područje Europe navodi se oko 30 vrsta, a u hrvatskoj flori 15 vrsta i 1 podvrsta. Od kojih su za šumska staništa značajne vrste – hrvatska perunika (I. croatica Horvat et M. D. Horvat), germanska perunika (I. germanica L.), uskolisna perunika (I. graminea L.), sibirska perunika (I. sibirica L), ilirska perunika (I. illyrica L.), žuta perunika (I. pseudacorus L.), šarena perunika (I. variegata L.). Sadržaj 1. Uvod .................................................................................................................................. 1 1.1. Prirodno stanište ......................................................................................................... 1 1.2. Morfološke značajke.................................................................................................. -

COMPARATIVE MORPHOLOGICAL ANALYSES of the GENUS Iris L., Pallidae SERIES (A
NAT. CROAT. VOL. 8 No 4 369¿384 ZAGREB December 31, 1999 ISSN 1330-0520 original scientific paper / izvorni znanstveni rad . UDK 579.2 581.4(1-924.4/.5) COMPARATIVE MORPHOLOGICAL ANALYSES OF THE GENUS Iris L., Pallidae SERIES (A. KERN.) TRINAJSTI] (Iridaceae) BO@ENA MITI]1 &ZINKA PAVLETI] Department of Botany, Faculty of Science, University of Zagreb, Maruli}ev trg 20/II, HR–10 000 Zagreb Miti}, B. & Pavleti}, Z.: Comparative morphological analyses of the genus Iris L., Pallidae se- ries (A. Kern.) Trinajsti} (Iridaceae). Nat. Croat., Vol. 8, No. 4., 369–384, 1999, Zagreb. The qualitative morphological features of the Alpine-Dinaric and cultivated populations, as well as of the herbarium specimens of the genus Iris, series Pallidae, were investigated. The height of the plant, leaves, inflorescence, flowers (spathe, perigonium, androecium, gynoecium), fruits and seeds were analysed. It was established that the morphological features analysed did not show pheno- typic plasticity. The following qualitative morphological features have possible diagnostic signifi- cance: intensity of the leaf vein, spathe colour, perigonium colour, and pollen and seed morphol- ogy. Of no diagnostic use are ratio of filament and anther lengths (individual variability), and size and form of the fruits (intrapopulation variability). Key words: genus Iris L., series Pallidae (A. Kern.) Trinajsti}, Alpine-Dinaric region, morphology Miti}, B. & Pavleti}, Z.: Komparativna morfolo{ka analiza roda Iris L., serije Pallidae (A. Kern.) Trinajsti} (Iridaceae). Nat. Croat., Vol. 8, No. 4., 369–384, 1999, Zagreb. Istra`ene su kvalitativne morfolo{ke osobine alpsko-dinarskih i kultiviranih populacija, te her- bariziranih primjeraka roda Iris, serije Pallidae. -

Uzgoj Perunika Rod Iris
UZGOJ PERUNIKA ROD IRIS Šarlija, Ksenija Undergraduate thesis / Završni rad 2014 Degree Grantor / Ustanova koja je dodijelila akademski / stručni stupanj: Josip Juraj Strossmayer University of Osijek, Faculty of agriculture / Sveučilište Josipa Jurja Strossmayera u Osijeku, Poljoprivredni fakultet Permanent link / Trajna poveznica: https://urn.nsk.hr/urn:nbn:hr:151:110354 Rights / Prava: In copyright Download date / Datum preuzimanja: 2021-10-05 Repository / Repozitorij: Repository of the Faculty of Agrobiotechnical Sciences Osijek - Repository of the Faculty of Agrobiotechnical Sciences Osijek SVEUČILIŠTE JOSIPA JURJA STROSSMAYERA U OSIJEKU POLJOPRIVREDNI FAKULTET U OSIJEKU Ksenija Šarlija, apsolvent Prediplomski studij smjera Hortikultura UZGOJ PERUNIKA (ROD IRIS) Završni rad Osijek, 2014. SVEUČILIŠTE JOSIPA JURJA STROSSMAYERA U OSIJEKU POLJOPRIVREDNI FAKULTET U OSIJEKU Ksenija Šarlija, apsolvent Prediplomski studij smjera Hortikultura UZGOJ PERUNIKA (ROD IRIS) Završni rad Povjerenstvo za ocjenu i obranu završnog rada: 1. prof.dr.sc. Nada Parađiković, predsjednik 2. mag.ing. Monika Tkalec, mentor 3. doc.dr.sc. Tomislav Vinković, član Osijek, 2014. ZAHVALA Ovom prilikom se zahvaljujem Poljoprivrednom fakultetu u Osijeku i svim profesorima koji su mi tijekom perioda studiranja pomogli u savladanju gradiva i konačnog uspjeha. Posebno bih se zahvalila asistentici i mentorici Moniki Tkalec koja me puna razumijevanja u sve uputila. Zahvaljujem i prof.dr.sc. Nadi Parađiković, nositeljici modula Povrćarstvo i cvjećarstvo, čija me terenska nastava -

COMPARATIVE MORPHOLOGICAL ANALYSES of the GENUS Iris L., Pallidae SERIES (A
NAT. CROAT. VOL. 8 No 4 369¿384 ZAGREB December 31, 1999 ISSN 1330-0520 original scientific paper / izvorni znanstveni rad . UDK 579.2 581.4(1-924.4/.5) COMPARATIVE MORPHOLOGICAL ANALYSES OF THE GENUS Iris L., Pallidae SERIES (A. KERN.) TRINAJSTI] (Iridaceae) BO@ENA MITI]1 &ZINKA PAVLETI] Department of Botany, Faculty of Science, University of Zagreb, Maruli}ev trg 20/II, HR–10 000 Zagreb Miti}, B. & Pavleti}, Z.: Comparative morphological analyses of the genus Iris L., Pallidae se- ries (A. Kern.) Trinajsti} (Iridaceae). Nat. Croat., Vol. 8, No. 4., 369–384, 1999, Zagreb. The qualitative morphological features of the Alpine-Dinaric and cultivated populations, as well as of the herbarium specimens of the genus Iris, series Pallidae, were investigated. The height of the plant, leaves, inflorescence, flowers (spathe, perigonium, androecium, gynoecium), fruits and seeds were analysed. It was established that the morphological features analysed did not show pheno- typic plasticity. The following qualitative morphological features have possible diagnostic signifi- cance: intensity of the leaf vein, spathe colour, perigonium colour, and pollen and seed morphol- ogy. Of no diagnostic use are ratio of filament and anther lengths (individual variability), and size and form of the fruits (intrapopulation variability). Key words: genus Iris L., series Pallidae (A. Kern.) Trinajsti}, Alpine-Dinaric region, morphology Miti}, B. & Pavleti}, Z.: Komparativna morfolo{ka analiza roda Iris L., serije Pallidae (A. Kern.) Trinajsti} (Iridaceae). Nat. Croat., Vol. 8, No. 4., 369–384, 1999, Zagreb. Istra`ene su kvalitativne morfolo{ke osobine alpsko-dinarskih i kultiviranih populacija, te her- bariziranih primjeraka roda Iris, serije Pallidae. -

Taksonomski Status, Rasprostranjenost I Morfološke Značajke Autohtonih Vrsta Roda Iris L
Taksonomski status, rasprostranjenost i morfološke značajke autohtonih vrsta roda Iris L. (Iridaceae) u Hrvatskoj Žaček, Josip Undergraduate thesis / Završni rad 2015 Degree Grantor / Ustanova koja je dodijelila akademski / stručni stupanj: University of Zagreb, Faculty of Forestry / Sveučilište u Zagrebu, Šumarski fakultet Permanent link / Trajna poveznica: https://urn.nsk.hr/urn:nbn:hr:108:556111 Rights / Prava: In copyright Download date / Datum preuzimanja: 2021-09-29 Repository / Repozitorij: University of Zagreb Faculty of Forestry and Wood Technology ŠUMARSKI FAKULTET PREDDIPLOMSKI STUDIJ ŠUMARSTVO JOSIP ŽAČEK TAKSONOMSKI STATUS, RASPROSTRANJENOST I MORFOLOŠKE ZNAČAJKE AUTOHTONIH VRSTA RODA IRIS L. U HRVATSKOJ ZAVRŠNI RAD ZAGREB, rujan 2015. Zavod: Zavod za šumarsku genetiku, dendrologiju i botaniku Predmet: Šumarska botanika Mentor: Prof. dr. sc. Jozo Franjić Asistent: Dr. sc. Daniel Krstonošić Student: Josip Žaček JMBAG: 0068210787 Akad. godina: 2014/2015 Mjesto,datum Zagreb, 25. rujna 2015. godine obrane: Sadržaj rada: Slika: 18 Tablica: 0 Navoda literature: 10 Sažetak: Rod Iris L. je rasprostranjen na Sjevernoj polutci. Rod se odlikuje velikim brojem vrsta, podvrsta, varijeteta i križanaca. Za područje Europe navodi se oko 30 vrsta, a u hrvatskoj flori 15 vrsta i 1 podvrsta. Od kojih su za šumska staništa značajne vrste – hrvatska perunika (I. croatica Horvat et M. D. Horvat), germanska perunika (I. germanica L.), uskolisna perunika (I. graminea L.), sibirska perunika (I. sibirica L), ilirska perunika (I. illyrica L.), žuta -

Gap Analysis Final Report
University of Primorska Science and Research Centre of Koper Institute for Biodiversity Studies WWF Project Reference No 9Z1387.05 “Protected Areas for a Living Planet – Dinaric Arc Ecoregion Project” Protected Area Gap Analysis (Final Report) Peter Glasnovi ć, BSc Boris Krystufek, PhD Andrej Sovinc, MSc Mileta Bojovi ć, BSc Deni Porej, PhD December 2009 WWF Dinaric Arc Ecoregion Project Protected Area Gap Analysis The Final Report by: University of Primorska Science and Research Centre of Koper Institute for Biodiversity Studies Garibaldijeva 1 6000 Koper Tel.: ++386 5 663 77 00, fax: ++386 5 663 77 10 E-mail: [email protected] Regional Scientific Coordinator: Peter Glasnovi ć, BSc; Boris Krystufek, PhD; Andrej Sovinc, MSc Cartography: Mileta Bojovi ć, BSc National Scientific Coordinators: Leon Kebe, BSc (Slovenia); Irina Zupan, MSc (Croatia); Senka Barudanovi ć, PhD (Bosnia and Herzegovina); Dragan Roganovi ć, PhD (Montenegro); Genti Kromidha, PhD (Albania) External experts: Boris Sket, PhD; Maja Zagmaister, PhD; Borut Štumberger, BSc WWF Mediterranean Programme Office: Director of Conservation Deni Porej, PhD Project Leader Stella Šatali ć, MSc Partners of the project: TNC (The Nature Conservancy), EuroNatur, Institute for Nature Conservation in Albania (Albania), University of Sarajevo – Faculty of Science (Bosnia and Herzegovina), State Institute for Nature Protection (Croatia), Institute for Nature Protection (Montenegro) 2 WWF Dinaric Arc Ecoregion Project Protected Area Gap Analysis Acknowledgments: Dragan Kova čevi ć, Banja Luka -

A New Spontaneous Hybrid Between the Cultivated and Wild Iris Species from Serbia
Bulletin of the Natural History Museum, 2018, 11: 189-210. Received 04 Sep 2018; Accepted 20 Nov 2018. doi:10.5937/bnhmb1811189N UDC: 582.711.714(497.11) Original scientific paper A NEW SPONTANEOUS HYBRID BETWEEN THE CULTIVATED AND WILD IRIS SPECIES FROM SERBIA MARJAN NIKETIĆ1*, GORDANA TOMOVIĆ2, SONJA SILJAK-YAKOVLEV3 1 Natural History Museum, 11000 Belgrade, Serbia, e-mail: [email protected] 2 Institute of Botany and Botanical Garden, Faculty of Biology, University of Belgrade, 11000 Belgrade, Serbia, e-mail: [email protected] 3 Ecologie Systématique Evolution, Univ. Paris-Sud, CNRS, AgroParisTech, Université Paris-Saclay, 91400 Orsay, France, e-mail: [email protected] A new spontaneous wilding hybrid, Iris ×seminaturalis Niketić, Tomović & Šiljak-Yak. (I. ×germanica L. s.l. × I. reichenbachii Heuff.) from C Serbia was described and illustrated. The comparative analyses of the morphological cha- racters and genome sizes were performed on I. ×seminaturalis and its parents. A new taxonomic review of the complex I. ×germanica s.l. was also presented. Key words: vascular flora, Iris ×seminaturalis, Iris ×germanica, spontaneous hybrid, Serbia INTRODUCTION With six subgenera, 301 currently accepted species, 17 hybrids and 27 additional non-type subspecies (Barker & Govaerts 2018), genus Iris L. has 190 NIKETIĆ, M. ET AL.: A NEW SPONTANEOUS IRIS HYBRID FROM SERBIA the richest taxonomic diversity in the family Iridaceae (Asparagales). Five subgenera are widespread in the Palaearctic, while subgenus Limniris (Tausch) Spach is distributed in the Holarctic. Most of the European species belong to the type subgenus with rhizomatous stocks and equitant ensiform leaves (Webb & Chater 1980). Representatives of the type section, I. -

Atti Del Museo Di Storia Di Trieste
ATTI DEL MUSEOMUSEO CCIVICOIVICO DIDI STORSTORIAIA NATURALENATURALE DIDI TRTRIESTEIESTE naturale A I stor I D CO I V I C MUSEO DEL DEL I ATT TRIESTE 2014 VOVOL.L. 59 56 - -2018 2013 ATTI DEL MUSEO CIVICO DI STORIA NATURALE ATTI DI TRIESTE DEL MUSEO CIVICO DI STORIA NATURALE DI TRIESTE VOL. 59 - 2018 ISSN: 0335-1576 DIRIGENTE DIRETTORE RESPONSABILE DEL PERIODICO Laura Carlini Fanfogna COMITATO SCIENTIFICO Deborah Arbulla, paleontologia Pietro Brandmayr, entomologia Nicola Bressi, zoologia TRIESTEAndrea 2014 Colla, entomologia VOL. 56 - 2013 Guido Ferilli, botanica Pier Luigi Nimis, botanica REDAZIONE Livio Fogar con Gianni Pistrini Museo Civico di Storia Naturale via Tominz, 4 – 34139 Trieste – Italia Tel.: +39406758227/662 – Fax: +390406758230 E-mail: [email protected]; [email protected] www.retecivica.trieste.it/triestecultura/musei In copertina: Podarcis muralis ♂ Tolmezzo (foto di Gianluca Rassati) On the cover: Podarcis muralis ♂ Tolmezzo (photo of Gianluca Rassati) Finito di stampare nel mese di dicembre 2018 da Lithostampa ISSN: 0335-1576 ATTI DEL MUSEO CIVICO DI STORIA NATURALE DI TRIESTE VOL. 59 - 2018 TRIESTE 2018 Atti Mus. Civ. St. Nat. Trieste 59 2018 5/20 XII 2018 ISSN: 0335-1576 LA VISIONE DEI MINERALI DEL MUSEO CIVICO DI STORIA NATURALE DI TRIESTE ENRICO FRANGIPANI Via dei Fabbri, n. 1, 34124 Trieste – E-mail: [email protected] Abstract – The vision of the minerals of Trieste Natural History Museum. Among the many objectives of an exhi- bition, there is not only the valorization of the specimen but also the possibility of finding correlations among different disciplines and thus contribute to a dissemination of solid scientific knowledge. -
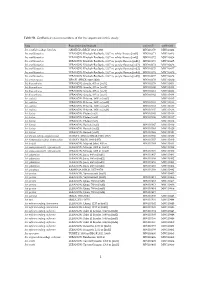
Table S1. Genbank Accession Numbers of the Iris Sequenced in This Study
Table S1. GenBank accession numbers of the Iris sequenced in this study. Taxa Population [individual] trnL-trnF matK-trnK Iris acutiloba subsp. lineolata ARMENIA (RBGK 2012-1109) MW110370 MW110422 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, white flowers [ind1] MW110371 MW110423 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, white flowers [ind2] MW110372 MW110424 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, purple flowers [ind1] MW110373 MW110425 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, purple flowers [ind2] MW110374 MW110426 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, purple flowers [ind3] MW110375 MW110427 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, purple flowers [ind4] MW110376 MW110428 Iris antilibanotica LEBANON: Kheibeh-Baalbeck, 1337 m, purple flowers [ind5] MW110377 MW110429 Iris atropurpurea ISRAEL (RBGK 1998-2808) MW110378 MW110430 Iris bismarkiana LEBANON: Sarada, 435 m [ind1] MW110379 MW110431 Iris bismarkiana LEBANON: Sarada, 435 m [ind2] MW110380 MW110432 Iris bismarkiana LEBANON: Sarada, 435 m [ind3] MW110381 MW110433 Iris bismarkiana LEBANON: Sarada, 435 m [ind4] MW110382 MW110434 Iris cedretii LEBANON: Bcharre, 1900 m [ind1] MW110435 Iris cedretii LEBANON: Bcharre, 1900 m [ind2] MW110383 MW110436 Iris cedretii LEBANON: Bcharre, 1900 m [ind3] MW110384 MW110437 Iris cedretii LEBANON: Bcharre, 1900 m [ind4] MW110385 MW110438 Iris histrio LEBANON: Ehden [ind1] MW110365 MW110416 Iris histrio LEBANON: Ehden [ind2] MW110366 MW110417 Iris histrio LEBANON: Ehden [ind3] MW110418 Iris histrio LEBANON: Barouk [ind1] MW110367 MW110419 Iris histrio LEBANON: Barouk [ind2] MW110368 MW110420 Iris histrio LEBANON: Barouk [ind3] MW110369 MW110421 Iris iberica subsp. elegantissima TURKEY: 2200 m (RBGK 1999-4347) MW110386 MW110439 Iris kirkwoodiae subsp. kirkwoodiae TURKEY (RBGK 1994-2407) MW110387 MW110440 Iris lortetii LEBANON: Mays el Jabal, 640 m MW110388 MW110441 Iris mesopotamica (I. -

NOM NOM Plantlist Lieu De Récolte Origine Hemerocallis Esculenta Koidzumi Hemerocallis Esculenta Koidz
NOM NOM Plantlist Lieu de récolte Origine Hemerocallis esculenta Koidzumi Hemerocallis esculenta Koidz. Xanthorrhoeaceae Iles Sakkhalines J.B. St. Petersbourg (Rs) Hemerocallis flava M Hotta. Hemerocallis lilioasphodelus L. Liliaceae Japon Hemerocallis middendorfii. Hemerocallis middendorffii Trautv. & C.A.Mey. Liliaceae Russie Suisse Hemerocallis middendorfii. Hemerocallis middendorffii Trautv. & C.A.Mey. Liliaceae Lyon Hemerocallis minor Hemerocallis minor Mill. Xanthorrhoeaceae Mt Chamar Daban (Rs) Mojmir Pavleka (Cz) Hemerocallis minor Hemerocallis minor Mill. Xanthorrhoeaceae Altaï Krai, 200 m (Rs) J.B. Osnabrûck (Ge) Iris acutiloba CA Mey. Iris acutiloba C.A.Mey. Iridaceae Iris albertii Regel. Iris albertii Regel. Iridaceae Iris albertii Regel. Iris albertii Regel. Iridaceae Ruffier Iris aphylla L. Iris aphylla L. Iridaceae Iris aphylla L. Iris aphylla L. Iridaceae Iris aphylla. Iris aphylla L. Iridaceae Iris aphylla. Iris aphylla L. Iridaceae Allemagne Iris arenaria W et K. Iris arenaria Waldst. & Kit. Iridaceae Iris atropurpurea Backer. Iris atropurpurea Backer. Iridaceae Iris balcana Janka. Iris reichenbachii Heuff. Iridaceae Iris barbata nana. Iris 'Barbata Nana' Iridaceae Lepage Iris bloudowi Ledeb. Iris bloudowi Ledeb. Iridaceae Kazakhstan Ruffier Iris bloudowi Ledeb. Iris bloudowi Ledeb. Iridaceae Sibérie Chitelet Iris bloudowii Ledeb. Iris bloudowii Ledeb. Iridaceae Iris bracteata S Watson. Iris bracteata S Watson. Iridaceae Iris brevicaulis Iris brevicaulis Raf. Iridaceae Iris bulleyana Iris bulleyana Dykes Iridaceae Cult. E. Lauroz Iris bulleyana Iris bulleyana Dykes Iridaceae Zheduo Shan, Sechuan (Sn) V. Holubec (Cz) Iris bulleyana Dykes Iris bulleyana Dykes Iridaceae Yunnan (Sn) J.B. Nancy (Ga) Iris caroliniana S Wats. Iris virginica L.. Iridaceae Iris caucasica Hoffmann. Iris caucasica Hoffmann. Iridaceae Iris caucasica. Iris caucasica Hoffmann. Iridaceae Iris chamaeiris Bertol. -

ACTA BOTANICA CROATICA Sadržaj Vol. 1—50* Contents of Volumes 1—50
Acta Bot. Croat. 50, 147—204, 1891. CODEN: ABCRA2 ISSN 0365-0588 ACTA BOTANICA CROATICA Sadržaj Vol. 1— 50* Contents of Volumes 1— 50 Vol. 1 (1925) Gjurašin, Stj.: Mycogalopsis retinospora nov. gen. et nov. spec, et son développement ........................................................................ 1—14 Horvat, I.: Razvoj i srodstvo paprati Cheilanthes (Development and Affinities of the Fern Cheilanthes)........................................... 15—21 Škorić, V.: Mikoriza u nekih Thymelaeacea (Die Mykorrhiza bei eini gen Thymelaeaceen) ..................................................................... 22—24 Juriste, P.: Die Geschwindigkeitsänderungen der Protoplasmaströ mung in der P flan zen zellen .......................................................... 25—31 Arnold, Z.: Peklova hipoteza o mikogenom podrijetlu aleurona (Peklo’s Hypothese über den mykogenen Ursprung des Aleu- rons) ................................................................................................. 32—36 Vouk, V.: Uber den plastidogenen Ursprung der Aleuronkörner . 37—43 Boić, D.: O kemijskom karakteru peridije, kapilieija i membrana spora u miksomiceta (Über den chemischen Charakter der Peridie, des Kapillitiums und der Sporenmembranen bei My xomyzeten) ................................................................................... 44—63 Ercegović, A.: Litofitska vegetacija vapnenaca i dolomita u Hrvat skoj (La végétation des lithophytes sur les calcaires et les dolo mites en C ro a tie )........................................................................... -
Medicinal and Aromatic Plants in Albania, Bosnia-Herzegovina, Bulgaria, Croatia and Romania
Wolfgang Kathe, Susanne Honnef & Andreas Heym Medicinal and Aromatic Plants in Albania, Bosnia-Herzegovina, Bulgaria, Croatia and Romania BfN – Skripten 91 Federal Agency for Nature Conservation 2003 Medicinal and Aromatic Plants in Albania, Bosnia-Herzegovina, Bulgaria, Croatia and Romania A study of the collection of and trade in medicinal and aromatic plants (MAPs), relevant legislation and the potential of MAP use for financing nature conservation and protected areas Wolfgang Kathe, Susanne Honnef & Andreas Heym (WWF Deutschland / TRAFFIC Europe-Germany) This study was carried out by WWF Deutschland and TRAFFIC Europe-Germany on behalf of the German Federal Agency for Nature Conservation (BfN), Bonn and Vilm. May 2003 Editors: Susanne Honnef, e-mail: [email protected] Wolfgang Kathe, e-mail: [email protected] Cover design and layout: Marion Blume Formatting: Karin Berkhoudt Publisher: Bundesamt für Naturschutz (BfN) (German Federal Agency for Nature Conservation) Konstantinstrasse 110, D-53179 Bonn, Germany Tel.: +49 228 / 8491 - 0 Fax: +49 228 / 8491 - 200 Internet: http://www.bfn.de BfN-Skripten are not available in book trade BfN-Skripten may be ordered from International Academy for Nature Conservation Isle of Vilm, D - 18581 Putbus, Germany e-mail: [email protected] download: www.bfn.de/09/090203.htm The publisher takes no guarantee for correctness, details and completeness of statements and views in this report as well as no guarantee for respecting private rights of third parties. Views expressed in the papers published in this issue of BfN-Skripten are those of the authors and do not necessarily represent those of the publisher. Printed on 100 % recycled paper by the printing office of the Federal Ministry for the Environment, Nature Conservation and Nuclear Safety.