Pest Risk Analysis for Lycorma Delicatula
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Antimicrobial and Antioxidant Activity of the Leaves, Bark and Stems of Liquidambar Styraciflua L
Int.J.Curr.Microbiol.App.Sci (2016) 5(1): 306-317 ISSN: 2319-7706 Volume 5 Number 1(2016) pp. 306-317 Journal homepage: http://www.ijcmas.com Original Research Article http://dx.doi.org/10.20546/ijcmas.2016.501.029 Antimicrobial and Antioxidant Activity of the Leaves, Bark and Stems of Liquidambar styraciflua L. (Altingiaceae) Graziele Francine Franco Mancarz1*, Ana Carolina Pareja Lobo1, Mariah Brandalise Baril1, Francisco de Assis Franco2 and Tomoe Nakashima1 1Pharmaceutical ScienceDepartment, Universidade Federal do Paraná, Curitiba, PR, Brazil 2Coodetec Desenvolvimento, Produção e Comercialização Agrícola Ltda, Cascavel, PR, Brazil *Corresponding author A B S T R A C T K e y w o r d s The genus Liquidambar L. is the best-known genus of the Altingiaceae Horan family, and species of this genus have long been used for the Liquidambar treatment of various diseases. Liquidambar styraciflua L., which is styraciflua, popularly known as sweet gum or alligator tree, is an aromatic deciduous antioxidant tree with leaves with 5-7 acute lobes and branched stems. In the present activity, study, we investigated the antimicrobial and antioxidant activity of aerial antimicrobial parts of L.styraciflua. Antimicrobial activity was evaluated using the activity, microdilution methodology. The DPPH and phosphomolybdenum methods microdilution method, were used to assess the antioxidant capacity of the samples. The extracts DPPH assay showed moderate or weak antimicrobial activity. The essential oil had the lowest MIC values and exhibited bactericidal action against Escherichia Article Info coli, Enterobacter aerogenes and Staphylococcus aureus. The ethyl acetate fraction and the butanol fraction from the bark and stem showed the best Accepted: antioxidant activity. -

What Is a Tree in the Mediterranean Basin Hotspot? a Critical Analysis
Médail et al. Forest Ecosystems (2019) 6:17 https://doi.org/10.1186/s40663-019-0170-6 RESEARCH Open Access What is a tree in the Mediterranean Basin hotspot? A critical analysis Frédéric Médail1* , Anne-Christine Monnet1, Daniel Pavon1, Toni Nikolic2, Panayotis Dimopoulos3, Gianluigi Bacchetta4, Juan Arroyo5, Zoltán Barina6, Marwan Cheikh Albassatneh7, Gianniantonio Domina8, Bruno Fady9, Vlado Matevski10, Stephen Mifsud11 and Agathe Leriche1 Abstract Background: Tree species represent 20% of the vascular plant species worldwide and they play a crucial role in the global functioning of the biosphere. The Mediterranean Basin is one of the 36 world biodiversity hotspots, and it is estimated that forests covered 82% of the landscape before the first human impacts, thousands of years ago. However, the spatial distribution of the Mediterranean biodiversity is still imperfectly known, and a focus on tree species constitutes a key issue for understanding forest functioning and develop conservation strategies. Methods: We provide the first comprehensive checklist of all native tree taxa (species and subspecies) present in the Mediterranean-European region (from Portugal to Cyprus). We identified some cases of woody species difficult to categorize as trees that we further called “cryptic trees”. We collected the occurrences of tree taxa by “administrative regions”, i.e. country or large island, and by biogeographical provinces. We studied the species-area relationship, and evaluated the conservation issues for threatened taxa following IUCN criteria. Results: We identified 245 tree taxa that included 210 species and 35 subspecies, belonging to 33 families and 64 genera. It included 46 endemic tree taxa (30 species and 16 subspecies), mainly distributed within a single biogeographical unit. -

Synteny Analysis in Rosids with a Walnut Physical Map Reveals Slow Genome Evolution in Long-Lived Woody Perennials Ming-Cheng Luo1, Frank M
Luo et al. BMC Genomics (2015) 16:707 DOI 10.1186/s12864-015-1906-5 RESEARCH ARTICLE Open Access Synteny analysis in Rosids with a walnut physical map reveals slow genome evolution in long-lived woody perennials Ming-Cheng Luo1, Frank M. You2, Pingchuan Li2, Ji-Rui Wang1,4, Tingting Zhu1, Abhaya M. Dandekar1, Charles A. Leslie1, Mallikarjuna Aradhya3, Patrick E. McGuire1 and Jan Dvorak1* Abstract Background: Mutations often accompany DNA replication. Since there may be fewer cell cycles per year in the germlines of long-lived than short-lived angiosperms, the genomes of long-lived angiosperms may be diverging more slowly than those of short-lived angiosperms. Here we test this hypothesis. Results: We first constructed a genetic map for walnut, a woody perennial. All linkage groups were short, and recombination rates were greatly reduced in the centromeric regions. We then used the genetic map to construct a walnut bacterial artificial chromosome (BAC) clone-based physical map, which contained 15,203 exonic BAC-end sequences, and quantified with it synteny between the walnut genome and genomes of three long-lived woody perennials, Vitis vinifera, Populus trichocarpa,andMalus domestica, and three short-lived herbs, Cucumis sativus, Medicago truncatula, and Fragaria vesca. Each measure of synteny we used showed that the genomes of woody perennials were less diverged from the walnut genome than those of herbs. We also estimated the nucleotide substitution rate at silent codon positions in the walnut lineage. It was one-fifth and one-sixth of published nucleotide substitution rates in the Medicago and Arabidopsis lineages, respectively. We uncovered a whole-genome duplication in the walnut lineage, dated it to the neighborhood of the Cretaceous-Tertiary boundary, and allocated the 16 walnut chromosomes into eight homoeologous pairs. -

Stuttgarter Beiträge Zur Naturkunde Serie B (Geologie Und Paläontologie)
Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie) Herausgeber: Staatliches Museum für Naturkunde, Rosenstein 1, D-70191 Stuttgart Stuttgarter Beitr. Naturk. Ser. B Nr. 306 58 pp., 56 figs. Stuttgart, 31. 8. 2001 New parasitic wasps from Baltic amber (Insecta: Hymenoptera: Dryinidae) By Massimo Olmi, Viterbo and Günter Bechly, Stuttgart With 56 Figures Summary Four new species of the parasitic wasp family Dryinidae, Dryinus reifi n. sp., Dryinus muenchi n. sp., Dryinus wunderlichi n. sp., and Palaeodryinus groehni n. gen. n. sp., are de- scribed from Baltic amber. These new species belong to the widespread and diverse subfami- ly Dryininae. A neotype is designated for Harpactosphecion gracile (BRUES, 1933b) comb. nov. which was originally described in the extant genus Thaumatodryinus and is here trans- ferred to the fossil genus Harpactosphecion within Dryininae, just like Harpactosphecion? deletum (BRUES, 1933b) comb. nov. The spelling of the names of the species here attributed to the genus Harpactosphecion is changed to gracile, deletum, filicorne, and sucinum in accor- dance with Art. 30 IRZN. New specimens of Dryinus bruesi (OLMI, 1984), Dryinus janzeni OLMI, 1999b, Harpactosphecion filicorne (BRUES, 1923), and Palaeoanteon janzeni OLMI, 1999b are figured, and a new subfamily Palaeoantoninae n. subf. is established because of the new evidence. The fossil subfamily Laberitinae OLMI, 1989 is regarded as unavailable taxon since the type genus Laberites PONOMARENKO, 1988 is not available either. Updated keys to the subfamilies of Dryinidae, the genera of Dryininae, and all dryinid species from Baltic am- ber are provided, as well as an annotated catalogue of all fossil Dryinidae. Zusammenfassung Vier neue Arten der parasitischen Wespenfamilie Dryinidae, Dryinus reifi n. -

WO 2012/104728 Al 9 August 2012 (09.08.2012) P O P C T
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date WO 2012/104728 Al 9 August 2012 (09.08.2012) P O P C T (51) International Patent Classification: AO, AT, AU, AZ, BA, BB, BG, BH, BR, BW, BY, BZ, C07D 311/92 (2006.01) A61K 36/77 (2006.01) CA, CH, CL, CN, CO, CR, CU, CZ, DE, DK, DM, DO, A61K 31/352 (2006.01) C07D 493/08 (2006.01) DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, HN, HR, HU, ID, IL, IN, IS, JP, KE, KG, KM, KN, KP, KR, (21) International Application Number: KZ, LA, LC, LK, LR, LS, LT, LU, LY, MA, MD, ME, PCT/IB20 12/000372 MG, MK, MN, MW, MX, MY, MZ, NA, NG, NI, NO, NZ, (22) International Filing Date: OM, PE, PG, PH, PL, PT, QA, RO, RS, RU, RW, SC, SD, 1 February 2012 (01 .02.2012) SE, SG, SK, SL, SM, ST, SV, SY, TH, TJ, TM, TN, TR, TT, TZ, UA, UG, US, UZ, VC, VN, ZA, ZM, ZW. (25) Filing Language: English (84) Designated States (unless otherwise indicated, for every (26) Publication Language: English kind of regional protection available): ARIPO (BW, GH, (30) Priority Data: GM, KE, LR, LS, MW, MZ, NA, RW, SD, SL, SZ, TZ, 61/438,395 1 February 20 11 (01.02.201 1) US UG, ZM, ZW), Eurasian (AM, AZ, BY, KG, KZ, MD, RU, TJ, TM), European (AL, AT, BE, BG, CH, CY, CZ, DE, (71) Applicant (for all designated States except US): UNIVER¬ DK, EE, ES, FI, FR, GB, GR, HR, HU, IE, IS, IT, LT, LU, SITY OF THE WEST INDIES [JM/JM]; A Regional In LV, MC, MK, MT, NL, NO, PL, PT, RO, RS, SE, SI, SK, stitution Established, By Royal Charter, Mona Campus, SM, TR), OAPI (BF, BJ, CF, CG, CI, CM, GA, GN, GQ, Kingston 7 (JM). -

Lycorma Delicatula
Bulletin OEPP/EPPO Bulletin (2020) 50 (3), 477–483 ISSN 0250-8052. DOI: 10.1111/epp.12702 European and Mediterranean Plant Protection Organization Organisation Europe´enne et Me´diterrane´enne pour la Protection des Plantes PM 7/144 (1) Diagnostics PM 7/144 (1) Lycorma delicatula Specific scope Specific approval and amendment This Standard describes a diagnostic protocol for Lycorma Approved in 2020-08. delicatula.1 This Standard should be used in conjunction with PM 7/ 76 Use of EPPO diagnostic protocols. Their validity needs to be confirmed (T. Bourgoin, pers. 1. Introduction comm.). Lycorma delicatula (White, 1845) (spotted lanternfly) is a planthopper indigenous to China, Taiwan and Vietnam 2. Identity where it is not a major pest, but damage has been reported in forests on Ailanthus altissima (tree of heaven) and on Preferred name: Lycorma delicatula (White, 1845). various fruit trees such as Actinidia (kiwi, etc.), Malus (ap- Other names: Aphaena delicatula White, 1845, Lycorma ple, etc.), Prunus (plum, etc.). It has been introduced into delicatulum (White, 1845). the Republic of Korea, Japan and the United States of Taxonomic position: Hemiptera, Auchenorrhyncha, Fulgo- America where it is considered to be showing invasive ridae, Lycorma. behaviour and causing economic damage. Lycorma EPPO Code: LYCMDE. delicatula is a polyphagous pest that causes direct damage Phytosanitary categorization: EPPO A1 list. to plants by feeding on the phloem, with large numbers of individuals that may feed on the same plant. Direct damage 3. Detection is caused by sucking plant sap, and indirect damage by pro- ducing honeydew on which fungi and sooty moulds may 3.1. -

Effects of Habitat Fragmentation and Disturbance on Biodiversity and Ecosystem Functions
Effects of habitat fragmentation and disturbance on biodiversity and ecosystem functions Inauguraldissertation der Philosophisch-naturwissenschaftlichen Fakultät der Universität Bern vorgelegt von Christof Schüepp von Eschlikon TG Leiter der Arbeit: Prof. Dr. M. H. Entling Institut für Ökologie und Evolution | downloaded: 13.3.2017 Von der Philosophisch-naturwissenschaftlichen Fakultät angenommen. Bern, 14. Mai 2013 Der Dekan: Prof. Dr. S. Decurtins Originaldokument gespeichert auf dem Webserver der Universitätsbibliothek Bern Dieses Werk ist unter einem https://doi.org/10.7892/boris.54819 Creative Commons Namensnennung-Keine kommerzielle Nutzung-Keine Bearbeitung 2.5 Schweiz Lizenzvertrag lizenziert. Um die Lizenz anzusehen, gehen Sie bitte zu http://creativecommons.org/licenses/by-nc-nd/2.5/ch/ oder schicken Sie einen Brief an Creative Commons, 171 Second Street, Suite 300, San Francisco, California 94105, USA. source: Effects of habitat fragmentation and disturbance on biodiversity and ecosystem functions Creative Commons Licence Urheberrechtlicher Hinweis Dieses Dokument steht unter einer Lizenz der Creative Commons Namensnennung- Keine kommerzielle Nutzung-Keine Bearbeitung 2.5 Schweiz. http://creativecommons.org/licenses/by-nc-nd/2.5/ch/ Sie dürfen: dieses Werk vervielfältigen, verbreiten und öffentlich zugänglich machen Zu den folgenden Bedingungen: Namensnennung. Sie müssen den Namen des Autors/Rechteinhabers in der von ihm festgelegten Weise nennen (wodurch aber nicht der Eindruck entstehen darf, Sie oder die Nutzung des Werkes durch Sie würden entlohnt). Keine kommerzielle Nutzung. Dieses Werk darf nicht für kommerzielle Zwecke verwendet werden. Keine Bearbeitung. Dieses Werk darf nicht bearbeitet oder in anderer Weise verändert werden. Im Falle einer Verbreitung müssen Sie anderen die Lizenzbedingungen, unter welche dieses Werk fällt, mitteilen. Jede der vorgenannten Bedingungen kann aufgehoben werden, sofern Sie die Einwilligung des Rechteinhabers dazu erhalten. -

Characterization of the Complete Chloroplast Genome of Platycarya Strobilacea (Juglandaceae)
Conservation Genet Resour (2017) 9:79–81 DOI 10.1007/s12686-016-0624-x TECHNICAL NOTE Characterization of the complete chloroplast genome of Platycarya strobilacea (Juglandaceae) Jing Yan1 · Kai Han1 · Shuyun Zeng1 · Peng Zhao1 · Keith Woeste2 · Jianfang Li1 · Zhan-Lin Liu1 Received: 1 September 2016 / Accepted: 4 October 2016 / Published online: 6 October 2016 © Springer Science+Business Media Dordrecht 2016 Abstract. The whole chloroplast genome (cp genome) The overall AT content of the cp genome is 64 %, and the sequence of Platycarya strobilacea was characterized corresponding values of the LSC, SSC and IR regions are from Illumina pair-end sequencing data. The complete cp 66.4, 70.1 and 57.5 %, respectively. Phylogenetic analysis genome was 160,994 bp in length and contained a large confirmed the placement of P. strobilacea near to Juglans. single copy region (LSC) of 90,225 bp and a small single copy region (SSC) of 18,371 bp, which were separated Keywords Platycarya strobilacea · Hickory · China · by a pair of inverted repeat regions (IRs, 26,199 bp). The Conservation · Chloroplast genome genome contained 130 genes, including 85 protein-coding genes (80 PCG species), 36 tRNA genes (29 tRNA spe- cies) and 8 ribosomal RNA genes (4 rRNA species). Most Platycarya strobilacea, the only species in the monotypic genes occur as a single copy, but 15 genes are duplicated. genus Platycarya (Juglandaceae), mainly grows as scat- tered individuals in evergreen forests of South China (Chen et al. 2012). As one of main components of the local vegeta- tion, this species plays a key role in the forest ecosystems. -

Lycorma Delicatula (Hemiptera: Auchenorrhyncha: Fulgoridae: Aphaeninae) Finally, but Suddenly Arrived in Korea
Entomological Research 38 (2008) 281–286 RESEARCHBlackwell Publishing Ltd PAPER Lycorma delicatula (Hemiptera: Auchenorrhyncha: Fulgoridae: Aphaeninae) finally, but suddenly arrived in Korea Jung Min HAN1, Hyojoong KIM2, Eun Ji LIM1, Seunghwan LEE2, Yong-Jung KWON3 and Soowon CHO1 1 Department of Plant Medicine, Chungbuk National University, Cheongju, Korea 2 School of Agricultural Biotechnology, Seoul National University, Seoul, Korea 3 Division of Applied Biology and Chemistry, Kyungpook National University, Daegu, Korea Correspondence Abstract Soowon Cho, Department of Plant Medicine, Chungbuk National University, A history of name changes in two fulgorid species – Lycorma delicatula and Limois Cheongju 361-763, Korea. emelianovi – is reviewed. Lycorma delicatula was once mistakenly reported to Email: [email protected] occur in Korea. Now, it has suddenly become common in western Korea, creating the suspicion that it has recently arrived from China and settled in Korea. A brief Received 6 April 2008; accepted 26 August morphological and biological description of L. delicatula is provided, and its 2008. original Korean name, “ggot-mae-mi”, is revalidated. Limois emelianovi, sometimes considered a synonym of emeljanovi, is the correct name for this species, doi: 10.1111/j.1748-5967.2008.00188.x as emeljanovi is simply another transliteration of the personal name Emelianov, Emeljanov or Emel’yanov. The name emelianovi stands correct based on the International Code of Zoological Nomenclature code 32.5.1, because there is no internal evidence of an inadvertent error, and an incorrect transliteration is not considered an inadvertent error. The cytochrome oxidase I (COI) barcoding regions of both species were sequenced and compared for future reference. -
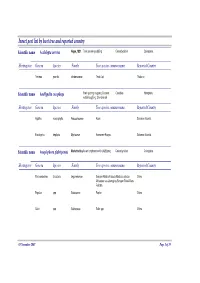
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae -

Reproduction and Potential Range Expansion of Walnut Twig Beetle Across the Juglandaceae
Biol Invasions (2018) 20:2141–2155 https://doi.org/10.1007/s10530-018-1692-5 ORIGINAL PAPER Reproduction and potential range expansion of walnut twig beetle across the Juglandaceae Andrea R. Hefty . Brian H. Aukema . Robert C. Venette . Mark V. Coggeshall . James R. McKenna . Steven J. Seybold Received: 10 June 2017 / Accepted: 19 February 2018 / Published online: 1 March 2018 Ó This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply 2018 Abstract Biological invasions by insects that vector this insect has expanded its geographic range by plant pathogens have altered the composition of colonizing naı¨ve hosts. The objective of this study was natural and urban forests. Thousand cankers disease to characterize limits to, and variation within, the host is a new, recent example and is caused by the complex range of P. juglandis and infer the extent to which of walnut twig beetle, Pityophthorus juglandis, and hosts might constrain the geographic distribution of the fungus, Geosmithia morbida, on susceptible hosts, the insect. We examined colonization and reproduc- notably some Juglans spp. and Pterocarya spp. Host tion by P. juglandis in no-choice laboratory experi- colonization by P. juglandis may be particularly ments with 11 Juglans spp., one Pterocarya sp., and important for disease development, but the beetle’s two Carya spp. over 2 years and found that all but the host range is not known. In the United States and Italy, Carya spp. were hosts. Reproduction was generally greater on Juglans californica, J. hindsii, and J. nigra, than on J. -

Genomics: Cracking the Mysteries of Walnuts
Review Article Genomics: cracking the mysteries of walnuts Fei Chen1*#, Junhao Chen2*, Zhengjia Wang2, Jiawei Zhang1, Meigui Lin1, Liangsheng Zhang1# 1State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops; Key Laboratory of Genetics, Breeding and Multiple Utilization of Corps (Fujian Agriculture and Forestry University), Ministry of Education, Fujian Provincial Key Laboratory of Haixia Applied Plant Systems Biology, Fujian Agriculture and Forestry University, Fuzhou 350002, China 2State Key Laboratory of Subtropical Silviculture, School of Forestry and Biotechnology, Zhejiang Agriculture and Forestry University, Hangzhou, 311300, China *Co-first author #Co-corresponding authors: Fei Chen, E-mail: [email protected]; Liangsheng Zhang, E-mail: [email protected] Abstract The Juglans plants are economically important by providing nuts, wood, and garden trees. They also play an important ecological role by supplying food for wild insects and animals. The decoding of genome sequences has fundamental values for understanding the evolution of Juglans plants and molecules, and is also a prerequisite for molecular breeding. During the last three years, the rapid development of sequencing technology has made walnut research into the genome era. Here, we reviewed the progress of genome sequencing of six Juglans species, the resequencing of four Juglans populations, as well as the genome sequencing of the closely related species Pterocarpa stenoptera. The analysis of the J. regia genome uncovers a whole genome duplication event. Based on the molecular dating of the divergence time of six Juglans species, we proposed this whole genome duplication event was associated with the cretaceous-Palaeogene (K-Pg) boundary happened ~65 million-year ago. Genomic sequences also provide clear details for understanding the evolution and development of GGT and PPO genes involved in fruit development.