The Leading-Edge Vortex of Swift Wings
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cali Types of the Common Swift Apus Apus: Aduit Cali Given at the Nest
Avocetta W 17: 141-146 (1993) Cali types of the Common Swift Apus apus: aduIt cali given at the nest VrNCENT BRETAGNOLLE CEBC-CNRS,F-79360-Beauvoirsur Niort, France Abstract - Vocalizations of the Common Swifts were studi ed during two consecutive springs in southern France. I found that three cali types were given by the adults at the nest, and these are described quantitatively. Significant differences in the acoustic parameters of the calls are highlighted, as well as. a probable sexual dimorphism. This, together with the precise signification of the different cali types, rernam however to be critically assessed by playback experiments. Introduction France), where a large colony is easily accessible for field studies (for further details, see Gory, this Swifts (Order Apodiformes) are unusual among birds volume).A sample of 22 accessible nests was because many of their Iife history traits (i.e. longevity, selected, for which extensive data on breeding biology delayed maturity, large egg size) make them atypical and breeding success were also available dating from compared to similar-sized birds (Lack 1956, Gaillard 1978 (Gory et Jeantet 1986). The birds were tape et al. 1989), and convergent evolution of life histories recorded by Uher 4400 or Nagra IV at a speed of 19.5 with, e.g., long lived seabirds such as cmJs, on Agfa PE43 tapes, and with a Seinnheiser Procellariiformes, have been suggested repeatedly omnidirectional microphone MD421 placed within, or (e.g. Lack and Lack 1951, Malacarne et al. 1991). at the entrance of, the nesting cavity. Calls were Although investigations have been carried out on the analysed on a Real Time Spectrograph, which Common Swift Apus apus with regard to demography performs a fast Fourier transform (Richard 1991) at a (Lack 1956, Perrins 1971, Lebreton et al. -

Fastest Migration Highest
GO!” Everyone knows birds can fly. ET’S But not everyone knows that “L certain birds are really, really good at it. Meet a few of these champions of the skies. Flying Acby Ellen eLambeth; art sby Dave Clegg! Highest You don’t have to be a lightweight to fly high. Just look at a Ruppell’s griffon vulture (left). One was recorded flying at an altitude of 36,000 feet. That’s as high as passenger planes fly! In fact, it’s so high that you would pass out from lack of oxygen if you weren’t inside a plane. How does the vulture manage? It has Fastest (on the level) Swifts are birds that have that name for good special blood cells that make a small amount reason: They’re speedy! The swiftest bird using its own of oxygen go a long way. flapping-wing power is the common swift of Europe, Asia, and Africa (below). It’s been clocked at nearly 70 miles per hour. That’s the speed limit for cars on some highways. Vroom-vroom! Fastest (in a dive) Fastest Migration With gravity helping out, a bird can pick up extra speed. Imagine taking a trip of about 4,200 And no bird can go faster than a peregrine falcon in a dive miles. Sure, you could easily do it in an airplane. after prey (right). In fact, no other animal on Earth can go as But a great snipe (right) did it on the wing in just fast as a peregrine: more than 200 miles per hour! three and a half days! That means it averaged about 60 miles The prey, by the way, is usually another bird, per hour during its migration between northern which the peregrine strikes in mid-air with its balled-up Europe and central Africa. -

E:\Wwwzoo~2.Org\Zoospr~1
Present distribution of Asian Pied Starlings S.K. Sharma Avifauna of ZSI Campus, Jodhpur C. Peruman et al. Sharma, I.K. (1988). Some birds around Pushkar Lake. Newsletter for Table 1. List of birds recorded in the Desert Regional Station, Birdwatchers 28(11&12): 7. ZSI. Sharma, A.K. and R. Singh (1993). Sighting of Green Barbet and nesting of Pied Myna at Jaipur. Newsletter for Bird Watchers 33(3): 53- Common name Scientific name Status* 54. Sharma, S.K. (2001). Impact of Indira Gandhi Canal on the desert Family: Phalacrocoracidae avifauna of Rajasthan. Report submitted to the Ministry of Environment Little Cormorant Phalacrocorax niger R and Forests, GOI, New Delhi. 459 pp. Vyas, R. (1992). Checklist of the birds of Kota district in South-East Family: Anatidae Rajsthan. Newsletter for Bird Watchers 32(11&12): 8-10. Spot-billed Duck Anas poecilorhyncha R Whistler, H. (1938). The ornithological survey of Jodhpur State. Journal of the Bombay Natural History Society 40: 213-235. Family: Accipitridae Black Kite Milvus migrans R Egyptian Vulture Neophron percnopterus R ACKNOWLEDGEMENT Indian White-backed Vulture Gyps bengalensis R This study was undertaken as a part of Dr. Salim Ali National Wildlife Tawny Eagle Aquila rapax R Shikra Accipiter trivirgatus R Fellwoship (1997) from the Ministry of Environment and Forests, New Delhi. The author is indebted to Dr. A.R. Rahmani, Director BNHS Family: Ardeidae Mumbai, Sh. R.G. Soni, PCCF, Rajasthan for guidance and constant Cattle Egret Bubulcus ibis R inspiration. Thanks are also due to Sh. Balveer -

White-Collared Swift: New to Ontario and Canada
77 White-collared Swift: New to Ontario and Canada Tristan ap Rheinallt On 10 June 2002, a White-collared sun rose, it soon warmed up and Swift (Streptoprocne zonaris) was became a bright, though somewhat seen briefly from the Marsh Trail at hazy, morning. I spent three very Rondeau Provincial Park. The pleasant hours pottering along the record, which has been accepted by first part of the trail, enjoying the the Ontario Bird Records sight of old friends such as Indigo Committee (Crins 2003), becomes Bunting (Passerina cyanea), Rose the first for Ontario and Canada. breasted Grosbeak (Pheucticus ludovicianus), Northern Cardinal Circumstances (Cardinalis cardinalis) and Orchard On the afternoon of 9 June 2002, I Oriole (Icterus spurius). I was arrived in Toronto from the UK at enjoying myself immensely and, for the start of a week-long birding trip. once, rarities were the last thing on It was my first visit to Canada since my mind. 1987 and my first to North America At 0800h, as I stood listening to since 1993. Although I knew that the birdsong and debating whether spring migration would be more or or not to attempt a photograph of a less over, there were several species superbly lit male Red-winged of breeding warblers that I hoped to Blackbird (Agelaius phoeniceus) add to my life list. One of these, perched on a nearby branch, I Prothonotary Warbler (Protonotaria noticed what appeared to be a swift citrea), was the reason I decided to flying directly towards me from the make Rondeau my first destination. direction of the lake. -

A Photographic Field Guide to the Birds of India
© Copyright, Princeton University Press. No part of this book may be 4 BIRDS OF INDIAdistributed, posted, or reproduced in any form by digital or mechanical means without prior written permission of the publisher. INTRODUCTION Birds of the Indian subcontinent an overview by Carol and Tim Inskipp The Indian subcontinent has a great wealth of birds, making it a paradise for the birdwatcher. The classic Handbook of the Birds of India and Pakistan by Salim Ali and S Dillon Ripley, which covers the whole region and was first published in 1968-1975, lists over 1,200 species. With additional recording and following the more up-to-date nomenclature in the Howard and Moore Complete Checklist of Birds of the World edited by E. C. Dickinson (2003) the current species total for the subcontinent stands at 1,375 species – 13 per cent of the world’s birds. A further relevant reference is Birds of South Asia: the Ripley Guide by Pamela Rasmussen and John Anderton, which has adopted much narrower species Dr. Salim Ali limits and, consequently, the latest edition (2012) recognises 1451 species in the region. Note that less than 800 species are found in all of North America. The Indian subcontinent is species-rich partly because of its wide altitudinal range extending from sea level up to the summit of the Himalayas, the world’s highest mountains. Another reason is the region’s highly varied climate and associated diverse vegetation. The extremes range from the almost rainless Great Indian or Thar Desert, where temperatures reach over 55°C, to the wet evergreen forests of the Assam Hills where 1,300 cm of rain a year have bee recorded at Cherrapunji – one of the wettest places on Earth, and the Arctic conditions of the Himalayan peaks where only alpine flowers and cushion plants flourish at over 4,900 m. -

Gliding for a Free Lunch: Biomechanics of Foraging Flight in Common Swifts (Apus Apus) Tyson L
© 2018. Published by The Company of Biologists Ltd | Journal of Experimental Biology (2018) 221, jeb186270. doi:10.1242/jeb.186270 RESEARCH ARTICLE Gliding for a free lunch: biomechanics of foraging flight in common swifts (Apus apus) Tyson L. Hedrick1,*,Cécile Pichot2 and Emmanuel de Margerie2,* ABSTRACT temporal (5–100 Hz) resolution, even for species where use of an Although the biomechanics of animal flight have been well studied in on-board satellite-based positioning package is currently infeasible laboratory apparatus such as wind tunnels for many years, the because of weight or other restrictions. Application of video applicability of these data to natural flight behaviour has been measurement methods has already revealed flight speeds well examined in few instances and mostly in the context of long-distance beyond those achieved in wind tunnels by quantifying mating or migration. Here, we used rotational stereo-videography to record the display behaviour in common swifts (Henningsson et al., 2010) and free-flight trajectories of foraging common swifts. We found that, hummingbirds (Clark, 2009), along with high-speed turns and despite their exquisite manoeuvring capabilities, the swifts only rarely pursuits in cliff swallows (Shelton et al., 2014). Field trajectory performed high-acceleration turns. More surprisingly, we also found recordings also revealed energy extraction from the ground to air that despite feeding on tiny insects probably moving with ambient wind speed gradient by foraging barn swallows (Warrick et al., flow, -

'Eastern' Common Swift Apus Apus Pekinensis in the Lakshadweep
AJU & SREENATH: ‘Eastern’ Common Swift 143 ‘Eastern’ Common Swift Apus apus pekinensis in the Lakshadweep Archipelago, with identification notes on juvenile birds Aju K. R. & Sreenath K. R. Aju K. R. & Sreenath K. R., 2019. ‘Eastern’ Common Swift Apus apus pekinensis in the Lakshadweep Archipelago, with identification notes on juvenile birds. Indian BIRDS 15 (5): 143–144. Aju K. R., Marine Biodiversity Division, Central Marine Fisheries Research Institute, Kochi 682018, Kerala, India. E-mail: [email protected] [Corresponding author.] [AKR] Sreenath K. R., Marine Biodiversity Division, Central Marine Fisheries Research Institute, Kochi 682018, Kerala, India. [SKR] Manuscript received on 05 November 2019. large, dark swift (Apodidae) was observed multiple times at Chetlat Island, Lakshadweep, in September 2019. It was A identified as a juvenile ‘Eastern’ Common SwiftApus apus pekinensis, which is an addition to the avifauna of the archipelago. We present a comprehensive discussion on its morphological characteristics, and its similarities and differences with the closely related Pallid Swift A. pallidus. a b Observation Chetlat Island (11.70ºN, 72.71ºE) is a sparsely populated atoll in 198a,b. Photographs taken in the evening, 24 September, 2019 (5.30 PM) the northern part of the Lakshadweep Archipelago. As part of a research tour, AKR reached the Island on 24 September 2019. On the same evening, he went birding along the eastern shore of the island and spotted a swift amongst a group of Barn Swallows Hirundo rustica and Sand Martins Riparia sp., on the northern end of the island. Despite being on the lookout for more, only a single individual of the swift was noted in this flock. -

The Common Swift Apus Apus—A New Bird for Sri Lanka
Correspondence 195 appearance and size, and its double-pointed casque with black false&gp=false&ev=Z&mr=1-12&bmo=1&emo=12&yr=all&byr=1900&eyr=2020. rim in front. It was feeding on a Ficus tree. Soon a female Great [Accessed on 05 August 2020.] Hornbill was also seen on the same tree [206]. They stayed there eBird. 2020b: Siberian Rubythroat Calliope calliope. Website URL: https://ebird.org/ map/sibrub?neg=true&env.minX=&env.minY=&env.maxX=&env.maxY=&zh=fa for few minutes and then flew away. The species was not seen lse&gp=false&ev=Z&mr=1-12&bmo=1&emo=12&yr=all&byr=1900&eyr=2020. on the subsequent visits to the national park. [Accessed on 05 August 2020.] eBird. 2020c. Rufous Woodpecker Micropternus brachyurus. Website URL: https://ebird.org/map/rufwoo2?neg=true&env.minX=&env. minY=&env.maxX=&env.maxY=&zh=false&gp=false&ev=Z&mr=1- 12&bmo=1&emo=12&yr=all&byr=1900&eyr=2020. [Accessed on 05 August 2020.] eBird. 2020d. Great Hornbill Buceros bicornis. Website URL: https://ebird.org/map/ grehor1?neg=true&env.minX=&env.minY=&env.maxX=&env.maxY=&zh=false&gp =false&ev=Z&mr=1-12&bmo=1&emo=12&yr=all&byr=1900&eyr=2020 [Accessed on 05 August 2020.] Grimmett, R., Inskipp, C., & Inskipp, T., 1998. Birds of the Indian Subcontinent. 1st ed. London: Christopher Helm, A & C Black. Pp. 1–888. Grimmett, R., Inskipp, C., & Inskipp, T., 2011. Birds of the Indian Subcontinent. 2nd ed. -
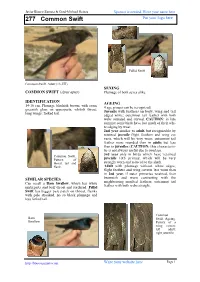
277 Common Swift Put Your Logo Here
Javier Blasco-Zumeta & Gerd-Michael Heinze Sponsor is needed. Write your name here Put your logo here 277 Common Swift Pallid Swift Common Swift. Adult (13-VII). SEXING COMMON SWIFT (Apus apus ) Plumage of both sexes alike. IDENTIFICATION AGEING 14-16 cm. Plumage blackish brown; with some 4 age groups can be recognized: greenish gloss on upperparts; whitish throat; Juvenile with feathers on body, wing and tail long wings; forked tail. edged white; outermost tail feather with both webs rounded and curved. CAUTION: in late summer some birds have lost much of their whi- te edging by wear. 2nd year similar to adult , but recognizable by retained juvenile flight feathers and wing co- verts, which will be very worn; outermost tail feather more rounded than in adults but less than in juveniles (CAUTION: this characteris- tic is not always useful due to overlap). 3rd year only in birds which have retained Common Swift. Pattern of juvenile 10th primay, which will be very throat, tail and strongly worn and reduced to the shaft. flank. Adult with plumage without white edges; flight feathers and wing coverts less worn than in 2nd year ; if outer primaries retained, then SIMILAR SPECIES brownish and worn contrasting with the Can recall a Barn Swallow , which has white neighbouring moulted feathers; outermost tail underparts and buff throat and forehead. Pallid feather with both webs straight. Swift has bigger pale patch on throat, flanks with pale streaked, no so black plumage and less forked tail. Common Barn Swift. Ageing. Swallow Pattern of a wing covert: left adult; right juvenile. -

The Coolest Bird: a Natural History of the Black Swift and Those Who
The Coolest Bird A Natural History of the Black Swift and Those Who Have Pursued It Rich Levad ~ 2007 ~ © 2010 American Birding Association. ~ Table of Contents ~ Foreward . .4 Acknowledgements . .6 1. Hawk Creek Falls, Colorado: A glimpse of things to come. 8 2. Semiahoo Bay, Washington: A new bird . 11 3. California’s Santa Cruz Coast: The first nest . 14 4. Johnston Canyon, Alberta: First inland nest site . 19 5. California: Charles and Enid Michaels at Yosemite . 22 6. California: Emily Smith and Berry Creek Falls . 27 7. California: Sequoia & King’s Canyon National Parks & San Jacinto Mountains . 31 8. Colorado: Niagara and Cataract Gulches . 34 9. Colorado: Al Knorr—more and more . 39 10. Arizona . 47 11. New Mexico . 53 12. Utah . 56 13. Southern California . 60 14. Northern Rocky Mountains: Montana, Idaho, Alberta . 67 15. NW Pacific Coast: British Columbia, Washington, Oregon . 73 16. Colorado post-Knorr: 1958-1996. 80 17. Colorado: Sue Hirshman and Box Canyon Falls . 87 18. Colorado 1995-1997 . 90 19. Colorado 1998 . 95 20. Colorado 1999-2000 . 102 21. Colorado: 2001-2002 . 107 22. Colorado and New Mexico 2003-2006 . 116 23. Following through in the Southern Rockies . 120 24. Recent events in the North . 125 25. The Southerners . 132 26. Today and Tomorrow . 136 Conservation Issues . 142 Bibliography . 149 3 ~ Foreword ~ y husband, Rich, was somewhat of a late comer to the hobby of bird watching; it was ducks that first lured him. He hunted them. In Colorado heavy fines can be levied for possessing certain Mspecies of ducks, so it pays to know the difference. -

First Record of Pallid Swift Apus Pallidus in Denmark and of Ssp
First record of Pallid Swift Apus pallidus in Denmark and of ssp. illyricus in northern Europe KASPER THORUP (Med et dansk resumé: Første danske fund af Gråsejler Apus pallidus illyricus) Examining skins of Common Swifts Apus apus Common Swift. According to Lewington (1999) and Pallid Swifts Apus pallidus at the Zoological this is because in Common Swift the outermost Museum, University of Copenhagen (ZMUC), primary (p10) is the longest while in Pallid Swift I discovered an aberrant swift in the series of p9 is longest. However, specimens of both Pallid Danish specimens that appeared identical to two and Common Swift have a highly variable wing specimens of the Pallid Swift subspecies illyricus formula, with most having the second outermost from Croatia in the same collection. Subsequent primary longest (p9 longest 86%, p10 longest measuring confirmed the identification. According 14%, N = 35; own measurements, ZMUC). Of the to the label the specimen was from Kirke Helsinge, two Croatian specimens of Pallid Swift, one had West Zealand 11 March 1993 and had been the outermost primary longest, the other having collected by Keld Bennike. The collection manager, the second outermost longest. Jan Bolding Kristensen (who prepared the · The wing length is 166 mm and the tail length 63 specimen), confirmed that the data were correctly mm, both falling within the range of copied from the original label, and that the bird both Common Swift and Pallid Swift (Chantler could not have been mixed up with birds from other & Driessens 1995). countries. The collector could, without remember- ing the specific circumstances, confirm the Danish Plumage origin. -

Colonial Breeding in the Barn Swallow (Hirundo Rustica) and Its Adaptive Significance
COLONIAL BREEDING IN THE BARN SWALLOW (HIRUNDO RUSTICA) AND ITS ADAPTIVE SIGNIFICANCE BARBARA DENNISTON SNAPP Social patterns in the swallow family (Hirun- whether active interactions among the breed- dinidae) involve varying degrees of gre- ing pairs allow the development of a Barn gariousness during the breeding season. Some Swallow colony, or whether such aggregations species, such as the Bank Swallow (Riparia are the result of a passive congregation at a TipaTia) and the Cliff Swallow (Petrochelidon nesting site. The presence or absence of such pyrrhonota), breed in dense colonies of up interactions may be inferred from an exami- to several hundred pairs. Other species, like nation of the variation in within-colony breed- the Tree Swallow (ITidopTocne bicoloT) and ing synchrony and occurrence of second- the Rough-winged Swallow (Stelgidopteryx brooding, as well as by direct observations of Tuficollis), are solitary breeders. A third pat- the social behavior involved in predator de- tern of social behavior is that of breeding in fense, foraging, and nest site selection. Last, small colonies of variable size. The Barn Swal- I attempted to relate the observations for Barn low (HiTundo mcstica) is such a species, found Swallows to those for other swallow species in nesting alone and in aggregations of 2 to 40 order to explain the rise of coloniality in the or more pairs ( Bent 1942). family as a whole. This diversity of social breeding habits is the result of adaptation to a number of en- METHODS viromnental pressures ( Crook 1965). These pressures may involve habitat and food type, I conducted the present investigation in Tompkins County, New York during the breeding seasons of dispersion of food supplies, and availability 1970 and 1971.