Leaf Morphological Differentiation Between Quercus Robur and Quercus Petraea Is Stable Across Western European Mixed Oak Stands Antoine Kremer, Jean Dupouey, J
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Species List
1 of 16 Claypits 20/09/2021 species list Group Taxon Common Name Earliest Latest Records acarine Aceria macrorhyncha 2012 2012 1 acarine Aceria nalepai 2018 2018 1 amphibian Bufo bufo Common Toad 2001 2018 6 amphibian Lissotriton helveticus Palmate Newt 2001 2018 5 amphibian Lissotriton vulgaris Smooth Newt 2001 2001 1 annelid Hirudinea Leech 2011 2011 1 bird Acanthis cabaret Lesser Redpoll 2013 2013 1 bird Acrocephalus schoenobaenus Sedge Warbler 2001 2011 2 bird Aegithalos caudatus Long-tailed Tit 2011 2014 2 bird Alcedo atthis Kingfisher 2020 2020 1 bird Anas platyrhynchos Mallard 2013 2018 4 bird Anser Goose 2011 2011 1 bird Ardea cinerea Grey Heron 2013 2013 1 bird Aythya fuligula Tufted Duck 2013 2014 1 bird Buteo buteo Buzzard 2013 2014 2 bird Carduelis carduelis Goldfinch 2011 2014 5 bird Chloris chloris Greenfinch 2011 2014 6 bird Chroicocephalus ridibundus Black-headed Gull 2014 2014 1 bird Coloeus monedula Jackdaw 2011 2013 2 bird Columba livia Feral Pigeon 2014 2014 1 bird Columba palumbus Woodpigeon 2011 2018 8 bird Corvus corax Raven 2020 2020 1 bird Corvus corone Carrion Crow 2011 2014 5 bird Curruca communis Whitethroat 2011 2014 4 bird Cyanistes caeruleus Blue Tit 2011 2014 6 bird Cygnus olor Mute Swan 2013 2014 4 bird Delichon urbicum House Martin 2011 2011 1 bird Emberiza schoeniclus Reed Bunting 2013 2014 2 bird Erithacus rubecula Robin 2011 2014 7 bird Falco peregrinus Peregrine 2013 2013 1 bird Falco tinnunculus Kestrel 2010 2020 3 bird Fringilla coelebs Chaffinch 2011 2014 7 bird Gallinula chloropus Moorhen 2013 -

Phytophthora Ramorum Sudden Oak Death Pathogen
NAME OF SPECIES: Phytophthora ramorum Sudden Oak Death pathogen Synonyms: Common Name: Sudden Oak Death pathogen A. CURRENT STATUS AND DISTRIBUTION I. In Wisconsin? 1. YES NO X 2. Abundance: 3. Geographic Range: 4. Habitat Invaded: 5. Historical Status and Rate of Spread in Wisconsin: 6. Proportion of potential range occupied: II. Invasive in Similar Climate YES NO X Zones United States: In 14 coastal California Counties and in Curry County, Oregon. In nursery in Washington. Canada: Nursery in British Columbia. Europe: Germany, the Netherlands, the United Kingdom, Poland, Spain, France, Belgium, and Sweden. III. Invasive in Similar Habitat YES X NO Types IV. Habitat Affected 1. Habitat affected: this disease thrives in cool, wet climates including areas in coastal California within the fog belt or in low- lying forested areas along stream beds and other bodies of water. Oaks associated with understory species that are susceptible to foliar infections are at higher risk of becoming infected. 2. Host plants: Forty-five hosts are regulated for this disease. These hosts have been found naturally infected by P. ramorum and have had Koch’s postulates completed, reviewed and accepted. Approximately fifty-nine species are associated with Phytophthora ramorum. These species are found naturally infected; P. ramorum has been cultured or detected with PCR but Koch’s postulates have not been completed or documented and reviewed. Northern red oak (Quercus rubra) is considered an associated host. See end of document for complete list of plant hosts. National Risk Model and Map shows susceptible forest types in the mid-Atlantic region of the United States. -

Quercus Robur (Fagaceae)
Quercus robur (Fagaceae) The map description EEBIO area The integrated map shows the distribution and changes in the areal’s boundaries of pedunculate oak (Quercus robur). Q. robur is the dominant forest- formative species in the belt of broadleaf and mixed 4 needleleaf-broadleaf forests in the plains of the European part of the former USSR (Sokolov et al. 1977). In the northern part of its areal Q. robur grows in river valleys. In the central part, it forms mixed forests with Picea abies; closer to the south – a belt of broadleaf forests where Q. robur dominates. At the areal’s south boundary it forms small (marginal) forests in ravines and flood-plains (Atlas of Areals and Resources… , 1976). Q. robur belongs to the thermophilic species. The low temperature bound of possible occurrence of oak forests is marked by an average annual of 2?C l (http://www.forest.ru – in Russian). Therefore, l l l l l l hypotretically, oak areal boundaries will shift along l ll l l l l l l l with the changes in the average annual temperature. l l l l l l l l l l l For Yearly map of averaged mean annual air l l l l l l l l l l l l l l l l l l lll temperature (Afonin A., Lipiyaynen K., Tsepelev V., l l l l l l l ll ll l l l 2005) see http://www.agroatlas.spb.ru Climate. l l l l l l l l l l l l l l l l Oak forests are of great importance for the water l l l l l ll l lll l l l l l l l l l l l l l l l l l l l regime and soil structure, especially on the steep l l l l l l l l l l l l l l l l l l l l l l l l l l l l l slopes of river valleys and in forest-poor areas. -

5 Fagaceae Trees
CHAPTER 5 5 Fagaceae Trees Antoine Kremerl, Manuela Casasoli2,Teresa ~arreneche~,Catherine Bod6n2s1, Paul Sisco4,Thomas ~ubisiak~,Marta Scalfi6, Stefano Leonardi6,Erica ~akker~,Joukje ~uiteveld', Jeanne ~omero-Seversong, Kathiravetpillai Arumuganathanlo, Jeremy ~eror~',Caroline scotti-~aintagne", Guy Roussell, Maria Evangelista Bertocchil, Christian kxerl2,Ilga porth13, Fred ~ebard'~,Catherine clark15, John carlson16, Christophe Plomionl, Hans-Peter Koelewijn8, and Fiorella villani17 UMR Biodiversiti Genes & Communautis, INRA, 69 Route d'Arcachon, 33612 Cestas, France, e-mail: [email protected] Dipartimento di Biologia Vegetale, Universita "La Sapienza", Piazza A. Moro 5,00185 Rome, Italy Unite de Recherche sur les Especes Fruitikres et la Vigne, INRA, 71 Avenue Edouard Bourlaux, 33883 Villenave d'Ornon, France The American Chestnut Foundation, One Oak Plaza, Suite 308 Asheville, NC 28801, USA Southern Institute of Forest Genetics, USDA-Forest Service, 23332 Highway 67, Saucier, MS 39574-9344, USA Dipartimento di Scienze Ambientali, Universitk di Parma, Parco Area delle Scienze 1lIA, 43100 Parma, Italy Department of Ecology and Evolution, University of Chicago, 5801 South Ellis Avenue, Chicago, IL 60637, USA Alterra Wageningen UR, Centre for Ecosystem Studies, P.O. Box 47,6700 AA Wageningen, The Netherlands Department of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556, USA lo Flow Cytometry and Imaging Core Laboratory, Benaroya Research Institute at Virginia Mason, 1201 Ninth Avenue, Seattle, WA 98101, -

Diagnosis of Phytophthora Ramorum in Trees
Diagnosis of PhytophthoraPhytophthora ramorumramorum in Trees by Dr. Kim D. Coder, Warnell School of Forest Resources, University of Georgia April, 2004 The organism shown to be responsible for SOD (sudden oak death) is Phytophthora ramorum, a fungus / yeast-like brown algae. This pathogen generates a number of symptoms in the trees infected. Of the trees and large shrubs shown to be infectable with this pathogen, some species have more serious stem and branch lesions like oaks, while other species have primarily leaf and twig lesions. In a single landscape, multiple hosts can keep the pathogen present for further attacks. This publication was prepared by reviewing approximately 35 research or disease announcement publications in Europe and North America. In addition, a number of factsheets and synthesized informa- tion guides were reviewed for continuity. This publication is designed for field diagnosis of SOD-like symptoms and related symptom sets on community trees. This publication should not be used in tree nursery situations, and is not a pathogen centered review. It is critical to seek pathological expertise and testing for confirming disease organism presence. A selected bibliography is available entitled “Sudden Oak Death – SOD: Bibliography of Important Literature.” (Coder, Kim D. 2004. University of Georgia, Warnell School of Forest Resources outreach publication SFR04-1. 2pp.). Names The tree syndrome or symptom set which characterizes attack by Phytophthora ramorum has common names of “ramorum dieback,” “ramorum blight,” “ramorum twig blight,”“ramorum leaf blight,” “ramorum stem canker,” “blood spot disease,” or “sudden oak death” (SOD). Phytophthora ramorum, and the less virulent Phytophthora nemorosa and Phytophthora pseudosyringae are all relatively new pathogen species recovered from trees which show ramorum blight symptoms. -

Proceedings, 18Th Central Hardwood Forest Conference; 2012 March 26-28; Morgantown, WV; Gen
United States Department of Agriculture Proceedings Forest Service 18th Central Hardwood Northern Research Station Forest Conference General Technical Report NRS-P-117 Morgantown, WV March 26-28, 2012 This document is being published in electronic format only (Web and CD). Any corrections or additions will be posted to the Web site (www.nrs.fs.fed.us/pubs). Cover photo of a morel mushroom by Shawn T. Grushecky, West Virginia University, used with permission. The findings and conclusions of each article in this publication are those of the individual author(s) and do not necessarily represent the views of the U.S. Department of Agriculture or the Forest Service. All articles were received in digital format and were edited for uniform type and style. Each author is responsible for the accuracy and content of his or her paper. The use of trade, firm, or corporation names in this publication is for the information and convenience of the reader. Such use does not constitute an official endorsement or approval by the U.S. Department of Agriculture or the Forest Service of any product or service to the exclusion of others that may be suitable. This publication/database reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed CAUTION: here have been registered. All uses of pesticides must be registered PESTICIDES by appropriate State and/or Federal agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fish or other wildlife—if they are not handled or applied properly. -

Forestry Department Food and Agriculture Organization of the United Nations
Forestry Department Food and Agriculture Organization of the United Nations Forest Genetic Resources Working Papers State of Forest and Tree Genetic Resources in the Republic of Moldova by Dragos Postolache Botanical Garden (Institute) Academy of Sciences Chisinau, Republic of Moldova 2004 Forest Resources Development Service Working Paper FGR/64E Forest Resources Division FAO, Rome, Italy Disclaimer The Forest Genetic Resources Working Papers report on issues and activities related to the conservation, sustainable use and management of forest genetic resources. The purpose of these papers is to provide early information on on-going activities and programmes, and to stimulate discussion. The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Quantitative information regarding the status of forest resources has been compiled according to sources, methodologies and protocols identified and selected by the author, for assessing the diversity and status of genetic resources. For standardized methodologies and assessments on forest resources, please refer to FAO, 2003. State of the World’s Forests 2003 ; and to FAO, 2001. Global Forest Resources Assessment 2000 (FRA2000). FAO Forestry Paper No 140 . Information can also be found at the FAO-Forestry Internet site (http://www.fao.org/forestry/Forestry.asp ). Comments and feedback are welcome. For further information please contact: Pierre Sigaud, Forestry Officer (Forest Genetic Resources) Forest Resources Division Forestry Department FAO, Viale delle Terme di Caracalla 00100 Rome, Italy Fax: +39 06 570 55 137 [email protected] For quotation: Postolache, D. -

Quercus ×Coutinhoi Samp. Discovered in Australia Charlie Buttigieg
XXX International Oaks The Journal of the International Oak Society …the hybrid oak that time forgot, oak-rod baskets, pros and cons of grafting… Issue No. 25/ 2014 / ISSN 1941-2061 1 International Oaks The Journal of the International Oak Society … the hybrid oak that time forgot, oak-rod baskets, pros and cons of grafting… Issue No. 25/ 2014 / ISSN 1941-2061 International Oak Society Officers and Board of Directors 2012-2015 Officers President Béatrice Chassé (France) Vice-President Charles Snyers d’Attenhoven (Belgium) Secretary Gert Fortgens (The Netherlands) Treasurer James E. Hitz (USA) Board of Directors Editorial Committee Membership Director Chairman Emily Griswold (USA) Béatrice Chassé Tour Director Members Shaun Haddock (France) Roderick Cameron International Oaks Allen Coombes Editor Béatrice Chassé Shaun Haddock Co-Editor Allen Coombes (Mexico) Eike Jablonski (Luxemburg) Oak News & Notes Ryan Russell Editor Ryan Russell (USA) Charles Snyers d’Attenhoven International Editor Roderick Cameron (Uruguay) Website Administrator Charles Snyers d’Attenhoven For contributions to International Oaks contact Béatrice Chassé [email protected] or [email protected] 0033553621353 Les Pouyouleix 24800 St.-Jory-de-Chalais France Author’s guidelines for submissions can be found at http://www.internationaloaksociety.org/content/author-guidelines-journal-ios © 2014 International Oak Society Text, figures, and photographs © of individual authors and photographers. Graphic design: Marie-Paule Thuaud / www.lecentrecreatifducoin.com Photos. Cover: Charles Snyers d’Attenhoven (Quercus macrocalyx Hickel & A. Camus); p. 6: Charles Snyers d’Attenhoven (Q. oxyodon Miq.); p. 7: Béatrice Chassé (Q. acerifolia (E.J. Palmer) Stoynoff & W. J. Hess); p. 9: Eike Jablonski (Q. ithaburensis subsp. -

Diversity of Wisconsin Rosids
Diversity of Wisconsin Rosids . oaks, birches, evening primroses . a major group of the woody plants (trees/shrubs) present at your sites The Wind Pollinated Trees • Alternate leaved tree families • Wind pollinated with ament/catkin inflorescences • Nut fruits = 1 seeded, unilocular, indehiscent (example - acorn) *Juglandaceae - walnut family Well known family containing walnuts, hickories, and pecans Only 7 genera and ca. 50 species worldwide, with only 2 genera and 4 species in Wisconsin Carya ovata Juglans cinera shagbark hickory Butternut, white walnut *Juglandaceae - walnut family Leaves pinnately compound, alternate (walnuts have smallest leaflets at tip) Leaves often aromatic from resinous peltate glands; allelopathic to other plants Carya ovata Juglans cinera shagbark hickory Butternut, white walnut *Juglandaceae - walnut family The chambered pith in center of young stems in Juglans (walnuts) separates it from un- chambered pith in Carya (hickories) Juglans regia English walnut *Juglandaceae - walnut family Trees are monoecious Wind pollinated Female flower Male inflorescence Juglans nigra Black walnut *Juglandaceae - walnut family Male flowers apetalous and arranged in pendulous (drooping) catkins or aments on last year’s woody growth Calyx small; each flower with a bract CA 3-6 CO 0 A 3-∞ G 0 Juglans cinera Butternut, white walnut *Juglandaceae - walnut family Female flowers apetalous and terminal Calyx cup-shaped and persistant; 2 stigma feathery; bracted CA (4) CO 0 A 0 G (2-3) Juglans cinera Juglans nigra Butternut, white -
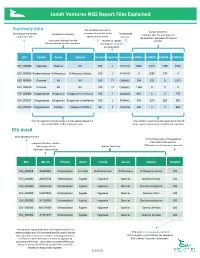
NGS Report Explained
Jonah Ventures NGS Report Files Explained Summary data The % of base pairs in the Sample identifiers Exact Sequence Variant sequence that match to the The detected Exact Sequence Variant Consensus Taxonomy The detected Numbers after the decimal point unique identifier species in the library. sequence. represent lab replicates of the same Taxonomic ranking from the Number of species sample. library matched to each sequence. matching the sequence at a given level. ESV Family Genus Species %match # Species Sequence S10001.1 S10002.1 S10003.1 S10003.2 ESV_000080 Fagaceae Quercus NA 100 7 AAAAAG… 3066 1340 5780 3462 ESV_000818 Orobanchaceae Orthocarpus Orthocarpus luteus 100 1 AAAAAG… 0 2582 249 0 ESV_000006 Poaceae NA NA 100 77 GAAAAG… 584 101 0 1039 ESV_000050 Poaceae NA NA 100 12 GAAAAG… 1328 0 0 0 ESV_000308 Polygonaceae Polygonum Polygonum humifusum 100 2 AAAAAG… 902 0 0 278 ESV_000027 Polygonaceae Eriogonum Eriogonum umbellatum 100 1 ATAAAG… 341 124 282 381 ESV_000520 Polygonaceae Fallopia Fallopia multiflora 99 1 AAAAAG… 126 0 0 994 “NA” at a taxonomic ranks means multiple species present in The number in each cell is the absolute number of the sample differ in that taxonomic rank times a given sequence was read by the sequencer. ESV detail Exact Sequence Variant The % of base pairs in the sequence that match to the species. Index identification number. that match to the species. Each sample has an Species Taxonomy 100% indicates all base pairs matched individual index number. ESV GB_ID Phylum Order Family Genus Species %match ESV_000818 -

Removal of Acorns of the Alien Oak Quercus Rubra on the Ground by Scatter-Hoarding Animals in Belgian Forests Nastasia R
B A S E Biotechnol. Agron. Soc. Environ. 2017 21(2), 127-130 Short note Removal of acorns of the alien oak Quercus rubra on the ground by scatter-hoarding animals in Belgian forests Nastasia R. Merceron (1, 2, 3), Aurélie De Langhe (3), Héloïse Dubois (3), Olivier Garin (3), Fanny Gerarts (3), Floriane Jacquemin (3), Bruno Balligand (3), Maureen Otjacques (3), Thibaut Sabbe (3), Maud Servranckx (3), Sarah Wautelet (3), Antoine Kremer (1), Annabel J. Porté (1, 2), Arnaud Monty (3) (1) BIOGECO, INRA, University of Bordeaux. FR-33610 Cestas (France). (2) BIOGECO, INRA, University of Bordeaux. FR-33615 Pessac (France). (3) University of Liège - Gembloux Agro-Bio Tech. Biodiversity and Landscape Unit. Passage des Déportés, 2. BE-5030 Gembloux (Belgium). E-mail: [email protected] Received on June 15, 2016; accepted on February 24, 2017. Description of the subject. Quercus rubra L. is considered an invasive species in several European countries. However, little is known about its dispersal in the introduced range. Objectives. We investigated the significance of animal dispersal of Q. rubra acorns on the ground by vertebrates in its introduced range, and identified the animal species involved. Method. During two consecutive autumns, the removal of acorns from Q. rubra and from a native oak was assessed weekly in forest sites in Belgium. We used automated detection camera traps to identify the animals that removed acorns. Results. Quercus rubra acorns were removed by wood mice (Apodemus sylvaticus L.), red squirrels (Sciurus vulgaris L.), rats (Rattus sp.), and wild boars (Sus scrofa L.). The two former are scatter-hoarding rodents and can be considered potential dispersers. -

Assessing Restoration Potential of Fragmented and Degraded Fagaceae Forests in Meghalaya, North-East India
Article Assessing Restoration Potential of Fragmented and Degraded Fagaceae Forests in Meghalaya, North-East India Prem Prakash Singh 1,2,* , Tamalika Chakraborty 3, Anna Dermann 4 , Florian Dermann 4, Dibyendu Adhikari 1, Purna B. Gurung 1, Saroj Kanta Barik 1,2, Jürgen Bauhus 4 , Fabian Ewald Fassnacht 5, Daniel C. Dey 6, Christine Rösch 7 and Somidh Saha 4,7,* 1 Department of Botany, North-Eastern Hill University, Shillong 793022, India; [email protected] (D.A.); [email protected] (P.B.G.); [email protected] (S.K.B.) 2 CSIR-National Botanical Research Institute, Council of Scientific & Industrial Research, Rana Pratap Marg, Lucknow 226001, Uttar Pradesh, India 3 Institute of Forest Ecosystems, Thünen Institute, Alfred-Möller-Str. 1, House number 41/42, D-16225 Eberswalde, Germany; [email protected] 4 Chair of Silviculture, University of Freiburg, Tennenbacherstr. 4, D-79085 Freiburg, Germany; anna-fl[email protected] (A.D.); fl[email protected] (F.D.); [email protected] (J.B.) 5 Institute for Geography and Geoecology, Karlsruhe Institute of Technology, Kaiserstr. 12, D-76131 Karlsruhe, Germany; [email protected] 6 Northern Research Station, USDA Forest Service, 202 Natural Resources Building, Columbia, MO 65211-7260, USA; [email protected] 7 Institute for Technology Assessment and Systems Analysis, Karlsruhe Institute of Technology, Karlstr. 11, D-76133 Karlsruhe, Germany; [email protected] * Correspondence: prem12fl[email protected] (P.P.S.); [email protected] (S.S.) Received: 5 August 2020; Accepted: 16 September 2020; Published: 19 September 2020 Abstract: The montane subtropical broad-leaved humid forests of Meghalaya (Northeast India) are highly diverse and situated at the transition zone between the Eastern Himalayas and Indo-Burma biodiversity hotspots.