An Analysis of the Microfauna and Microflora of The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Carnival Corporation Carnival Plc
UNITED STATES OMB APPROVAL SECURITIES AND EXCHANGE COMMISSION OMB Number: 3235-00595 Washington, D.C. 20549 Expires: February 28, 2006 SCHEDULE 14A Estimated average burden hours per response......... 12.75 Proxy Statement Pursuant to Section 14(a) of the Securities Exchange Act of 1934 (Amendment No. ) Filed by the Registrant x Filed by a Party other than the Registrant o Check the appropriate box: o Preliminary Proxy Statement o Confidential, for Use of the Commission Only (as permitted by Rule 14a-6(e)(2)) x Definitive Proxy Statement o Definitive Additional Materials o Soliciting Material Pursuant to Rule §240.14a-12 CARNIVAL CORPORATION CARNIVAL PLC (Name of Registrant as Specified In Its Charter) (Name of Person(s) Filing Proxy Statement, if other than the Registrant) Payment of Filing Fee (Check the appropriate box): x No fee required. o Fee computed on table below per Exchange Act Rules 14a-6(i)(1) and 0-11. 1. Title of each class of securities to which transaction applies: 2. Aggregate number of securities to which transaction applies: 3. Per unit price or other underlying value of transaction computed pursuant to Exchange Act Rule 0-11 (set forth the amount on which the filing fee is calculated and state how it was determined): 4. Proposed maximum aggregate value of transaction: 5. Total fee paid: SEC 1913 (03-04) Persons who are to respond to the Collection of information contained in this form are not required to respond unless the form displays a currently valid OMB cotrol number. o Fee paid previously with preliminary materials. -

Type, Figured and Cited Specimens in the Museum of Isle of Wight Geology (Isle of Wight, England)
THE GEOLOGICAL CURATOR VOLUME 6, No.5 CONTENTS PAPERS TYPE, FIGURED AND CITED SPECIMENS IN THE MUSEUM OF ISLE OF WIGHT GEOLOGY (ISLE OF WIGHT, ENGLAND). by J.D. Radley ........................................ ....................................................................................................................... 187 THE WORTHEN COLLECTION OF PALAEOZOIC VERTEBRATES AT THE ILLINOIS STATE MUSEUM by R.L. Leary and S. Turner ......................... ........................................................................................................195 LOST AND FOUND ......................................................................................................................................................207 DOOK REVIEWS ........................................................................................ ................................................................. 209 GEOLOGICAL CURATORS' GROUP .21ST ANNUAL GENERAL MEETING .................................................213 GEOLOGICAL CURATORS' GROUP -April 1996 TYPE, FIGURED AND CITED SPECIMENS IN THE MUSEUM OF ISLE OF WIGHT GEOLOGY (ISLE OF WIGHT, ENGLAND) by Jonathan D. Radley Radley, J.D. 1996. Type. Figured and Cited Specimens in the Museum of Isle of Wight Geology (Isle of Wight, England). The Geological Curator 6(5): 187-193. Type, figured and cited specimens in the Museum of Isle of Wight Geology are listed, as aconseouence of arecent collection survev and subseauentdocumentation work. Strengths.. currently lie in Palaeogene gas tropods, and Lower -

The Stratigraphical Framework for the Palaeogene Successions of the London Basin, UK
The stratigraphical framework for the Palaeogene successions of the London Basin, UK Open Report OR/12/004 BRITISH GEOLOGICAL SURVEY OPEN REPORT OR/12/004 The National Grid and other Ordnance Survey data are used The stratigraphical framework for with the permission of the Controller of Her Majesty’s Stationery Office. the Palaeogene successions of the Licence No: 100017897/2012. London Basin, UK Key words Stratigraphy; Palaeogene; southern England; London Basin; Montrose Group; Lambeth Group; Thames Group; D T Aldiss Bracklesham Group. Front cover Borehole core from Borehole 404T, Jubilee Line Extension, showing pedogenically altered clays of the Lower Mottled Clay of the Reading Formation and glauconitic sands of the Upnor Formation. The white bands are calcrete, which form hard bands in this part of the Lambeth Group (Section 3.2.2.2 of this report) BGS image P581688 Bibliographical reference ALDISS, D T. 2012. The stratigraphical framework for the Palaeogene successions of the London Basin, UK. British Geological Survey Open Report, OR/12/004. 94pp. Copyright in materials derived from the British Geological Survey’s work is owned by the Natural Environment Research Council (NERC) and/or the authority that commissioned the work. You may not copy or adapt this publication without first obtaining permission. Contact the BGS Intellectual Property Rights Section, British Geological Survey, Keyworth, e-mail [email protected]. You may quote extracts of a reasonable length without prior permission, provided a full acknowledgement is given of the source of the extract. Maps and diagrams in this book use topography based on Ordnance Survey mapping. © NERC 2012. -

Policy and Practice for the Protection Of
POLICY AND PRACTICE FOR THE PROTECTION OF GROUNDWATER SOUTHERN REGION APPENDIX W R A YtO @ 0^ ? CONTENTS 1. Introduction 2. Description of Southern Region 3. Office Locations and Administration 4. The Importance of Groundwater 1n Southern Region 5. Rainfall and Recharge 6. Geology and Hydrogeology 7. Use of National Policy with old Southern Region Maps until new maps and zones are produced 8. Cone1us1on/Summary Tables Table 1 Geological Succession 1n Southern Region Table 2 Classification of Strata 1n Southern Region Table 3 Correlation between old Southern Water Aquifer Protection Zone and new National Policy Protection Zones Figure 1 D1agrammat1c Comparison of old and new source protection zones Maps Map 1 Southern Region Area Map 2 Distribution of Rainfall and Public Water Supply Sources Map 3 Water Company Boundaries 1. INTRODUCTION Purpose of the Regional Appendix Th 1 s Reg1ona 1 Appendi x to the NRA " Pol 1 cy and Pract 1 ce for the Protection of Groundwater" provides Information specific to Southern Region. Details are given on the following subjects: * Description of Southern Region * Geology and Hydrogeology * Main Office Locations and Contacts relevant to groundwater matters * How to use the "Policy and Practice for the Protection of Groundwater" prior to the Introduction of new maps and protection zones Th1 s 1 s one of ten appendices that have been produced, each one specific to a different NRA region. Although the main document 1s a national one there are certain considerations, within the headings listed above, that are only relevant to this Region. Each appendix 1s produced to the same format with the necessary extra Information included. -

35. the "UPPER EOCENE, C.Om.P~'F/N~ T~F BARTON
Downloaded from http://jgslegacy.lyellcollection.org/ at University of California-San Diego on July 6, 2016 ~7~ MESSRS.GARDNER~ KEEPING, AND NONCKTON ON THE 35. The "UPPER EOCENE, C.om.p~'~F/n~ t~f BARTON ~nd UPPER BAGSHOT Fox,Ions. By J. S~ARXIE GARDSmZ, Esq., F.G.S., F.L.S., HE,mr KE~P~a, Esq., and H. W. MercaTor, Esq., F.G.S. (Read March 28, 1888.) THE introduction of the Oligocene stage into our classification has necessitated a partial revision of the grouping of our older British Tertiaries. Whether this introduction of a new primary division into the Tertiary system was necessary or expedient may still be ques- tioned ; but it has been generally adopted and is, for the time being, established. The division does not coincide in England with a marked change in either fauna or flora, though the series seems nevertheless tolerably complete and well developed; its limits, however widely stretched~ show that the Oligocene stage compares neither with the Eocene nor the Miocene in importance. Opinions have differed as to where the line of division should be drawn; whether this should be as low down as the top of the Barton Beds or at the base of the Headou Beds, or even higher. For our part, we think it desirable to uphold the view which places the demarcation between the top of the Bagshot Sands of Alum Bay and the base of the Lower Headon Series, though it is perfectly ob- vious that any such line in the midst of our series must be a purely artificial one. -

Ebook Download the Final Cut Ebook
THE FINAL CUT PDF, EPUB, EBOOK Michael Dobbs | 480 pages | 09 Apr 2010 | HarperCollins Publishers | 9780007375158 | English | London, United Kingdom The Final Cut ( film) - Wikipedia Trailers and Videos. Crazy Credits. Alternate Versions. Rate This. Episode Guide. In order to secure his retirement and establish Available on Amazon. Added to Watchlist. Top-Rated Episodes S1. Error: please try again. The Best Horror Movies on Netflix. Magnus wants to watch. DomProMedia - Series Complete. Use the HTML below. You must be a registered user to use the IMDb rating plugin. Episodes Seasons. Edit Cast Series cast summary: Ian Richardson Francis Urquhart 4 episodes, Diane Fletcher Elizabeth Urquhart 4 episodes, Paul Freeman Tom Makepeace 4 episodes, Isla Blair Claire Carlsen 4 episodes, Nickolas Grace Geoffrey Booza Pitt 4 episodes, Glyn Grain Rayner 4 episodes, Nick Brimble Corder 4 episodes, Dorothy Vernon Speaker 4 episodes, Andrew Seear Wolfin 4 episodes, Peter Symonds Polecutt 4 episodes, John Rowe Sir Clive Watling 3 episodes, Yolanda Vazquez Maria Passolides 3 episodes, Duggie Brown Joe Badger 3 episodes, Kevork Malikyan Nures 2 episodes, David Ryall Sir Bruce Bullerby 2 episodes, Joseph Long President Nicolaou 2 episodes, Cherith Mellor Hilary Makepeace 2 episodes, Erika Hoffman Princess 2 episodes, Leon Lissek Evanghelos Passolides 2 episodes, Tom Beasley Young King 2 episodes, Richard Bebb Political Commentator 2 episodes, Sue Edelson Newsreader 2 episodes, David Ashford Edit Storyline Francis Urquhart is too experienced a politician not to know that everything must end, even his long career as British prime minister. Genres: Drama. Edit Did You Know? Trivia Michael Dobbs , the author of the original book, was angered at the opening scene depicting Margaret Thatcher 's funeral. -

A Review of the Decapod Crustaceans from the Tertiary of the Isle of Wight, Hampshire, U
Bulletin of the Mizunami Fossil Museum, no. 38 (2012), p. 33–51, 4 pls., 2 figs., 2 tables. 33 © 202, Mizunami Fossil Museum A review of the decapod crustaceans from the Tertiary of the Isle of Wight, Hampshire, U. K, with description of three new species W. J. Quayle* and J. S. H. Collins** * 4, Argyll Street, Ryde, Isle of Wight, Hampsire, England<[email protected]> ** 8, Shaws Cottages, Perry Rise, London, SE23 2QN, and The Natural History Museum, London, SW7 5BD, England Abstract To the known decapod crustaceans of the Tertiary deposits of the Isle of Wight, Hampshire, the Early Eocene species, Basinotopus lamarkii (Desmarest) and Dromilites bucklandi Bell are introduced from the mainland, as are Glyphithyreus wetherelli (Bell), Rhachiosoma bispinosum Woodward, Coeloma (Litoricola) dentate (Woodward), Xanthilites bowerbanki Bell, and Xanthopsis unispinosa (M'Coy), the stratigrahic range of which has been extended to the London Clay on the Island. The development of an anomaly concerning Zanthopsis unispinosa, also present, if not peculiar to the island, is well established. A carapace of Harpactocarcinus sp. from this horizon is the first British record for the genus. Late Eocene species new to the island are Goniocypoda quaylei Crane, Orthakrolophus depressus? (Quayle and Collins) and Typilobus belli Quayle and Collins. Three new genera and species are described from the Late Eocene Headon Hill Formation; a callianassid, Vecticallichirus abditus; a goneplacid, Gonioplacoides minuta, and a hexapodid, Headonipus tuberculosus. New, superior material of the Late Eocene species, Typilobus obscurus Quayle and Collins, from the type locality, Colwell Bay necessitates a new description. A table of all known Tertiary species of decapod crustaceans from the Isle of Wight is appended. -
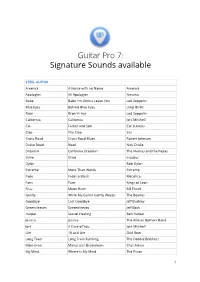
Signature Sounds Available
Guitar Pro 7: Signature Sounds available STEEL GUITAR America A Horse with no Name America Apologies All Apologies Nirvana Babe Babe I'm Gonna Leave You Led Zeppelin Blue Eyes Behind Blue Eyes Limp Bizkit Bron Bron Yr Aur Led Zeppelin California California Joni Mitchell Cat Father and Son Cat Stevens Clap The Clap Yes Cross Road Cross Road Blues Robert Johnson Drake Road Road Nick Drake Dreamin California Dreamin' The Mamas and the Papas Drive Drive Incubus Dylan Bob Dylan Extreme More Than Words Extreme Fade Fade to Black Metallica Fans Fans Kings of Leon Frizz Moon River Bill Frisell Gently While My Guitar Gently Weeps The Beatles Goodbye Last Goodbye Jeff Buckley Greensleeves Greensleeves Jeff Beck Harper Sexuel Healing Ben Harper Jessica Jessica The Allman Bothers Band Joni A Case of You Joni Mitchell Life 18 and Life Skid Row Long Train Long Train Running The Doobie Brothers Mainstreet Mainstreet Breakdown Chet Atkins My Mind Where Is My Mind The Pixies 1 No Rain No Rain Blind Melon November November Rain Guns 'N Roses Old Man Old Man Neil Young Overnight Overnight Bag Rory Gallagher Pinball Pinball Wizard The Who Presence Dear God Avenged Sevenfold Redemption Redemption Song Bob Marley Revolution Talkin 'Bout a Revolution Tracy Chapman Road Road Nick Drake September Ends Wake Me Up When September Ends Green Day Sixteen Tons Sixteen Tons Merle Travis Sky Eye Eye in the Sky Alan Pasons Project Sleep The Sleep Pantera Stairway Stairway to Heaven Led Zeppelin Stomp Bron-Y-Aur Stomp Led Zeppelin Summer All Summer Long Kid Rock Theater -

Port of Southampton Master Plan 2009-2030 Aerial Photograph of the Port of Southampton ABP Port of Southampton Master Plan 2009-2030 • Foreword
Port of Southampton Master Plan 2009-2030 Aerial photograph of the Port of Southampton ABP Port of Southampton Master Plan 2009-2030 • Foreword Foreword The Port of Southampton is the UK’s premier international gateway port. It connects trading nations across the world and helps the UK to benefit from the global economy. In 2007, we handled more cargo than ever before. In 2008, the Port broke another record with almost one million cruise passengers passing through the Port. It is, therefore, no surprise that the Port is recognised as one of very few UK international gateways whose commercial growth is critical to regional and national economic success. Doug Morrison Port Director, ABP Southampton Success for the Port is also success for Southampton and the community in the surrounding region. It is estimated that the Port supports around 10% of the City’s workforce directly and indirectly. We are also at the heart of the Solent maritime sector which is estimated to support up to economic conditions of 2009 in our forecasting as well as 77,000 jobs and be worth a massive £5.5 billion. identifying opportunities for the Port to contribute to a sustainable, low carbon economy. Our predecessors made far-sighted decisions to grow, consolidate and expand the Port. At the turn of the 20th When preparing the plan we have borne in mind three century they extended the Eastern Docks; in the 1920s and essentials: 30s they planned and built the Western Docks, and in the • the need to continue to attract investment and create 1960s they built one of the UK’s first container terminals. -

Barton-On-Sea Instability
Barton-on-Sea Instability Peter Ferguson NFDC Introduction •Geological setting •Engineering works •Ground movement & instability •Public safety & engagement •Management & maintenance •Monitoring •Ground investigations •Future schemes •Funding Location © Crown copyright and database rights 2014 Ordnance Survey 100026220 Geology Geology Geology Geology • Geology of Hampshire & Isle of Wight After Woodward 1904 Ian West 2000 Geology • Stratigraphy Plateau Gravel Bed L Pleistocene (fluvial deposits) Bed K Bed J Eocene Strata (Barton Sand Formation) Bed I Geology • Internationally important exposure • English Nature • Maintain exposures • SSSI rating • Unique exposure of Barton Beds Geology Geology Geology Translational Slides Engineering Works • 1930 – First Defences • Timber groynes & cliff drainage • 1939 to 1945 – Disrepair • 1950 Works become ineffective • Erosion rate 1-2m year • 1964 – 1968 Major works undertaken • Vertical piled revetment • Re-profiled slope • Sheet pile cut off wall • 1970’s • 5 rock groynes • 1990’s • Timber revetment replaced with rock revetment Engineering Works Ground Movement & Instability N • Landslide data Naish Barton-on-Sea Continuing mass movement 200m Ground Movement & Instability Cliff House Hotel Naish Farm Marine Drive W Barton Court Marine Drive E Ground Movement & Instability •Cliff House Hotel •Deep -seated failure • Occurred early 2001 • Displaced rock revetment at toe Cliff House Hotel Hosseyni et al. (In Press) Barton & Garvey (2011) Translational failure with mudslides Shear on base of D and possibly -

Track List: All Songs Written by Syd Barrett, Except Where Noted. ”UK Release” Side One 1
Track List: All songs written by Syd Barrett, except where noted. ”UK release” Side one 1. "Astronomy Domine" – 4:12 2. "Lucifer Sam" – 3:07 3. "Matilda Mother" – 3:08 4. "Flaming" – 2:46 5. "Pow R. Toc H." (Syd Barrett, Roger Waters, Rick Wright, Nick Mason) – 4:26 6. "Take Up Thy Stethoscope and Walk" (Roger Waters) – 3:05 Side two 1. "Interstellar Overdrive" (Syd Barrett, Roger Waters, Rick Wright, Nick Mason) – 9:41 2. "The Gnome" – 2:13 The Piper at the Gates of Dawn – August 5, 1967 3. "Chapter 24" – 3:42 Syd Barrett – guitar, lead vocals, back cover design 4. "The Scarecrow" – 2:11 5. "Bike" – 3:21 Roger Waters – bass guitar, vocals Rick Wright – Farfisa Compact Duo, Hammond organ, Certifications: piano, vocals TBR Nick Mason – drums, percussion "Astronomy Domine" Lime and limpid green, a second scene A fight between the blue you once knew. Floating down, the sound resounds Around the icy waters underground. Jupiter and Saturn, Oberon, Miranda and Titania. Neptune, Titan, Stars can frighten. Lime and limpid green, a second scene A fight between the blue you once knew. Floating down, the sound resounds Around the icy waters underground. Jupiter and Saturn, Oberon, Miranda and Titania. Neptune, Titan, Stars can frighten. Blinding signs flap, Flicker, flicker, flicker blam. Pow, pow. Stairway scare, Dan Dare, who's there? Lime and limpid green, the sounds around The icy waters under Lime and limpid green, the sounds around The icy waters underground. "Lucifer Sam" Lucifer Sam, siam cat. Always sitting by your side Always by your side. -

Pink Floyd, Revisited: Waters Doesn’T Disappoint by Christopher John Treacy, Boston Herald | September 9, 2006
9 Sep 2006 - Concert Review Boston Herald - Roger Waters Band Tweeter Center, Boston, MA, USA (8-Sep-06) Pink Floyd, revisited: Waters doesn’t disappoint By Christopher John Treacy, Boston Herald | September 9, 2006 No gimmicky light show or practiced cover band can match the majesty of seeing Roger Waters recreate his visionary work with Pink Floyd. And even though he’s currently touring under the guise of performing the legendary ‘Dark Side of The Moon’ that’s literally only the half of it. ‘Dark Side’ was originally performed as Act 2 of a Floyd show - exactly how Waters did it last night before a capacity crowd for the first of two gigs at Mansfield’s Tweeter Center. For all the anticipation surrounding Act 2, the first set was a much more compelling display. That’s not to knock Floyd’s 1973 masterpiece, which the musically muscled 11-piece band did commendable justice to. But hearing him lead his troops through crisp renditions of ‘Have a Cigar,’ ‘Sheep’ - during which an inflatable pink pig circled the venue – ‘Wish You Were Here,’ the opener ‘In the Flesh’ and ‘Shine on You Crazy Diamond (Parts 1-5),’ complete with multiple images of the late Syd Barrett flashing across the video screen, was infinitely more satisfying. During ‘Mother,’ when posing the question, ‘Should I trust the government?’ the crowd roared, but Waters disapprovingly shook his head as if to say, ‘You’ve missed the point.’ But there was no misinterpreting his messages of despair and disillusionment during the politically charged power-suite of ‘Southampton Dock,’ ‘The Fletcher Memorial Home’ and ‘Perfect Sense,’ the latter of which likens war to an arena sporting event.