Understanding Species-Microplastics Interactions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1 Creating a Species Inventory for a Marine Protected Area: the Missing
Katherine R. Rice NOAA Species Inventory Project Spring 2018 Creating a Species Inventory for a Marine Protected Area: The Missing Piece for Effective Ecosystem-Based Marine Management Katherine R. Rice ABSTRACT Over the past decade, ecosystem-based management has been incorporated into many marine- management administrations as a marine-conservation tool, driven with the objective to predict, evaluate and possibly mitigate the impacts of a warming and acidifying ocean, and a coastline increasingly subject to anthropogenic control. The NOAA Office of National Marine Sanctuaries (ONMS) is one such administration, and was instituted “to serve as the trustee for a network of 13 underwater parks encompassing more than 600,000 square miles of marine and Great Lakes waters from Washington state to the Florida Keys, and from Lake Huron to American Samoa” (NOAA, 2015). The management regimes for nearly all national marine sanctuaries, as well as other marine protected areas, have the goal of managing and maintaining biodiversity within the sanctuary. Yet none of those sanctuaries have an inventory of their known species nor a standardized protocol for measuring or monitoring species biodiversity. Here, I outline the steps required to compile a species inventory for an MPA, but also describe some of stumbling blocks that one might encounter along the way and offer suggestions on how to handle these issues (see Appendix A: Process for Developing the MBNMS Species Inventory (PD-MBNMS)). This project consists of three research objectives: 1. Determining what species inventory efforts exist, how they operate, and their advantages and disadvantages 2. Determining the process of creating a species inventory 3. -
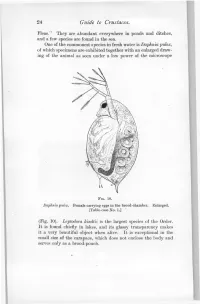
24 Guide to Crustacea
24 Guide to Crustacea. Fleas." They are abundant everywhere in ponds and ditches, and a few species are found in the sea. One of the commonest species in fresh water is Daphnia pulex, of which specimens are exhibited together with an enlarged draw- ing of the animal as seen under a low power of the microscope FIG. 10. Daphnia pulex. Female carrying eggs in the brood-chamber. Enlarged. [Table-case No. 1.] (Fig. 10). Leptodora kindtii is the largest species of the Order. It is found chiefly in lakes, and its glassy transparency makes it a very beautiful object when alive. It is exceptional in the small size of the carapace, which does not enclose the body and serves only as a brood-pouch. Ostracoda. 25 Sub-class II.—OSTRACODA. (Table-ease No. 1.) The number of somites, as indicated by the appendages, is smaller than in any other Crustacea, there being, at most, only two pairs of trunk-limbs behind those belonging to the head- region. The carapace forms a bivalved shell completely en- closing the body and limbs. There is a large, and often leg-like, palp on the mandible. The antennules and antennae are used for creeping or swimming. The Ostracoda (Fig. 10) are for the most part extremely minute animals, and only one or two of the larger species can be exhibited. They occur abundantly in fresh water and in FIG. 11. Shells of Ostracoda, seen from the side. A. Philomedes brenda (Myodocopa) ; B. Cypris fuscata (Podocopa); ('. Cythereis ornata (Podocopa): all much enlarged, n., Notch characteristic of the Myodocopa; e., the median eye ; a., mark of attachment of the muscle connecting the two valves of the shell. -

Biodiversity, Habitats, Flora and Fauna
1 North East inshore Biodiversity, Habitats, Flora and Fauna - Protected Sites and Species 2 North East offshore 3 East Inshore Baseline/issues: North West Plan Areas 10 11 Baseline/issues: North East Plan Areas 1 2 4 East Offshore (Please note that the figures in brackets refer to the SA scoping database. This is • SACs: There are two SACs in the plan area – the Berwickshire and North available on the MMO website) Northumberland Coast SAC, and the Flamborough Head SAC (Biodiv_334) 5 South East inshore • Special Areas of Conservation (SACs): There are five SACs in the plan area • The Southern North Sea pSAC for harbour porpoise (Phocoena phocoena) 6 South inshore – Solway Firth SAC, Drigg Coast SAC, Morecambe Bay SAC, Shell Flat and is currently undergoing public consultation (until 3 May 2016). Part of Lune Deep SAC and Dee Estuary SAC (Biodiv_372). The Sefton Coast the pSAC is in the offshore plan area. The pSAC stretches across the 7 South offshore SAC is a terrestrial site, mainly for designated for dune features. Although North East offshore, East inshore and offshore and South East plan areas not within the inshore marine plan area, the development of the marine plan (Biodiv_595) 8 South West inshore could affect the SAC (Biodiv_665) • SPAs: There are six SPAs in the plan area - Teesmouth and Cleveland 9 South west offshore • Special protection Areas (SPAs): There are eight SPAs in the plan area - Coast SPA, Coquet Island SPA, Lindisfarne SPA, St Abbs Head to Fast Dee Estuary SPA, Liverpool Bay SPA, Mersey Estuary SPA, Ribble and Castle SPA and the Farne Islands SPA, Flamborough Head and Bempton 10 North West inshore Alt Estuaries SPA, Mersey Narrows and North Wirral Foreshore SPA, Cliffs SPA (Biodiv_335) Morecambe Bay SPA, Duddon Estuary SPA and Upper Solway Flats and • The Northumberland Marine pSPA is currently undergoing public 11 North West offshore Marshes SPA (Biodiv_371) consultation (until 21 April 2016). -

Estudos Em Megabalanus Azoricus (Pilsbry, 1916)
Contributo para o conhecimento da biodiversidade regional – estudos em Megabalanus azoricus (Pilsbry, 1916). Dionísio, M.1, A. Pagarete2 & Costa, A.3 1, 2, 3 CIBIO – Centro de Investigação em Biodiversidade e Recursos Genéticos, Pólo Açores, Portugal; 2 – Departamento de Biologia, Universidade dos Açores, Rua da Mãe de Deus Apartado 1422, 9501-855 Ponta Delgada, Açores, Portugal. Tel. +351 296650100; Fax: +351 296650101 e-mail: 1 [email protected]; 2 [email protected]; 3 [email protected] RESUMO: Megabalanus azoricus (Pilsbry, 1916) é uma espécie nativa do arquipélago que apesar de ser reconhecida em toda a região onde sofre uma elevada exploração, só agora foi objecto de um estudo alargado. Apresentam-se aqui os primeiros resultados do projecto POCI/MAR/58185/2004 financiado pela FCT que visa o estudo dos aspectos da biologia desta espécie considerada pela OSPAR em risco e com necessidade de estudo urgente. A descrição de Megabalanus azoricus foi revista e pela primeira vez obtiveram-se larvas em laboratório. Iniciou-se uma inventariação de populações em várias ilhas do Arquipélago. Os estudos de reprodução realizados, baseados no estudo macroscópico e estereológico das gónadas de uma população de São Miguel permitiram verificar que estas cracas nos Açores apresentam-se sexualmente maturas ao longo de todo o ano, embora não se tenha registado produção de ovos em Novembro, Março e Agosto. Foram comparadas algumas populações provenientes de várias ilhas através da comparação das respectivas sequências de DNA de dois fragmentos COI-COII e 18S- ITS1 tendo-se verificado uma baixa variabilidade nos haplotipos de ambos os marcadores, o que não suporta níveis de diferenciação significativos entre as populações. -

Close Encounters
Environment International 140 (2020) 105792 Contents lists available at ScienceDirect Environment International journal homepage: www.elsevier.com/locate/envint Close encounters - microplastic availability to pelagic amphipods in sub- antarctic and antarctic surface waters T ⁎ Kirstie Jones-Williamsa,b, , Tamara Gallowayb, Matthew Colec, Gabriele Stowassera, Claire Waludaa, Clara Mannoa a British Antarctic Survey, High Cross, Madingley Road, Cambridge CB30ET, United Kingdom b University of Exeter, Streatham Campus, Northcote House, Exeter EX4 4QJ, United Kingdom c Plymouth Marine Laboratory, Prospect Place, Plymouth PL1 3DH, United Kingdom ARTICLE INFO ABSTRACT Handling Editor: Adrian Covaci This study investigated the distribution of plastic debris from the Atlantic portion of the Sub-Antarctic to the Keywords: Antarctic Peninsula. This region is home to some of the highest concentrations of zooplankton biomass but is also Microplastics threatened by increasing shipping traffic from fishing and the growing tourism market. Samples were collected Mespolastics using a surface-towed neuston net during the Austral summer 2018, aboard the RRS James Clark Ross. Using Encounter Rate Fourier Transform Infrared Spectrometry it was found that 45.6% of the plastic particles isolated from seawater Synthetic Fibres samples were sampling contamination, originating predominantly from the ship. Of the remaining particles, both Amphipods low density (polyethylene, polypropylene) and high-density (phenoxy and epoxy resins) polymers were found in Southern Ocean the surface water suggesting both long-range and local sources of origin. Whilst we found that micro and me- soplastic concentrations in seawater were significantly low (0.013 ± 0.005n/m3) compared to global averages, they were higher along the Antarctic Peninsula than the open ocean (Sub-Antarctic) stations. -

A Checklist of Turtle and Whale Barnacles
Journal of the Marine Biological Association of the United Kingdom, 2013, 93(1), 143–182. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S0025315412000847 A checklist of turtle and whale barnacles (Cirripedia: Thoracica: Coronuloidea) ryota hayashi1,2 1International Coastal Research Center, Atmosphere and Ocean Research Institute, The University of Tokyo, 5-1-5, Kashiwanoha, Kashiwa-shi, Chiba 277-8564 Japan, 2Marine Biology and Ecology Research Program, Extremobiosphere Research Center, Japan Agency for Marine–Earth Science and Technology A checklist of published records of coronuloid barnacles (Cirripedia: Thoracica: Coronuloidea) attached to marine vertebrates is presented, with 44 species (including 15 fossil species) belonging to 14 genera (including 3 fossil genera) and 3 families recorded. Also included is information on their geographical distribution and the hosts with which they occur. Keywords: checklist, turtle barnacles, whale barnacles, Chelonibiidae, Emersoniidae, Coronulidae, Platylepadidae, host and distribution Submitted 10 May 2012; accepted 16 May 2012; first published online 10 August 2012 INTRODUCTION Superorder THORACICA Darwin, 1854 Order SESSILIA Lamarck, 1818 In this paper, a checklist of barnacles of the superfamily Suborder BALANOMORPHA Pilsbry, 1916 Coronuloidea occurring on marine animals is presented. Superfamily CORONULOIDEA Newman & Ross, 1976 The systematic arrangement used herein follows Newman Family CHELONIBIIDAE Pilsbry, 1916 (1996) rather than Ross & Frick (2011) for reasons taken up in Hayashi (2012) in some detail. The present author Genus Chelonibia Leach, 1817 deems the subfamilies of the Cheonibiidae (Chelonibiinae, Chelonibia caretta (Spengler, 1790) Emersoniinae and Protochelonibiinae) proposed by Harzhauser et al. (2011), as well as those included of Ross & Lepas caretta Spengler, 1790: 185, plate 6, figure 5. -

Cirripedia of Madeira
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Universidade do Algarve Helgol Mar Res (2006) 60: 207–212 DOI 10.1007/s10152-006-0036-5 ORIGINAL ARTICLE Peter Wirtz Æ Ricardo Arau´jo Æ Alan J. Southward Cirripedia of Madeira Received: 13 September 2005 / Revised: 12 January 2006 / Accepted: 13 January 2006 / Published online: 3 February 2006 Ó Springer-Verlag and AWI 2006 Abstract We give a list of Cirripedia from Madeira phers. The marine invertebrates have been less studied Island and nearby deep water, based on specimens in and there has been no compilation of cirripede records the collection of the Museu Municipal do Funchal for Madeira, comparable to those for the Azores (Histo´ria Natural) (MMF), records mentioned in the archipelago (Young 1998a; Southward 1999). We here literature, and recent collections. Tesseropora atlantica summarize records from Madeira and nearby deep water Newman and Ross, 1976 is recorded from Madeira for and discuss their biogeographical implications. the first time. The Megabalanus of Madeira is M. az- oricus. There are 20 genera containing 27 species, of which 22 occur in depths less than 200 m. Of these Methods shallow water species, eight are wide-ranging oceanic forms that attach to other organisms or to floating The records are based on (1) the work of R.T. Lowe, objects, leaving just 13 truly benthic shallow water who sent specimens to Charles Darwin; (2) material in barnacles. This low diversity is probably a consequence the Museu Municipal do Funchal (Histo´ria Natural) of the distance from the continental coasts and the (MMF); (3) casual collecting carried out by residents or small area of the available habitat. -

Remarkable Convergent Evolution in Specialized Parasitic Thecostraca (Crustacea)
Remarkable convergent evolution in specialized parasitic Thecostraca (Crustacea) Pérez-Losada, Marcos; Høeg, Jens Thorvald; Crandall, Keith A Published in: BMC Biology DOI: 10.1186/1741-7007-7-15 Publication date: 2009 Document version Publisher's PDF, also known as Version of record Citation for published version (APA): Pérez-Losada, M., Høeg, J. T., & Crandall, K. A. (2009). Remarkable convergent evolution in specialized parasitic Thecostraca (Crustacea). BMC Biology, 7(15), 1-12. https://doi.org/10.1186/1741-7007-7-15 Download date: 25. Sep. 2021 BMC Biology BioMed Central Research article Open Access Remarkable convergent evolution in specialized parasitic Thecostraca (Crustacea) Marcos Pérez-Losada*1, JensTHøeg2 and Keith A Crandall3 Address: 1CIBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, Universidade do Porto, Campus Agrário de Vairão, Portugal, 2Comparative Zoology, Department of Biology, University of Copenhagen, Copenhagen, Denmark and 3Department of Biology and Monte L Bean Life Science Museum, Brigham Young University, Provo, Utah, USA Email: Marcos Pérez-Losada* - [email protected]; Jens T Høeg - [email protected]; Keith A Crandall - [email protected] * Corresponding author Published: 17 April 2009 Received: 10 December 2008 Accepted: 17 April 2009 BMC Biology 2009, 7:15 doi:10.1186/1741-7007-7-15 This article is available from: http://www.biomedcentral.com/1741-7007/7/15 © 2009 Pérez-Losada et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. -

Final Project Report
FINAL PROJECT REPORT Direct and indirect ecotoxicological impacts of microplastics on marine organisms Project acronym: PLASTOX Period covered: 01.01.2016 – 30.06.2019 Date of submission: 03.07.2019 Picture: Marco Capolupo (University of Bologna) Coordinating institute: SINTEF Ocean Project coordinator: Dr Andy Booth, Senior Researcher, Environmental Technology Dept., SINTEF Ocean Tel: +47 93089510 E-mail : [email protected] PLASTOX website address: https://www.sintef.no/projectweb/plastox/ CONSORTIUM Names Organisation Country SINTEF Ocean, Environmental Technology Department, Dr Andy Booth Norway Trondheim Dr Kaori Sakaguchi-Söder Technische Universität Darmstadt (TUDA), Darmstadt Germany Prof. Paula Sobral NOVA.ID FCT (NOVA-FCT), Caparica Portugal Prof. Laura Airoldi, Prof. Elena Alma Mater Studiorum - University of Bologna (UNIBO) Italy Fabbri, Prof. Giulio Zanaroli Aix-Marseille Université-Mediterranean Institute of Dr Richard Sempéré France Oceanography (AMU-MIO), Marseille Wageningen Marine Research (Wageningen University and Dr Jan Andries van Franeker Netherlands Research) (WUR), Den Helder Dr Tom Doyle, Dr Liam National University of Ireland Galway (NUIG), Galway Ireland Morrison Norwegian University of Science and Technology (NTNU) Dr Iurgi Salaberria Norway Trondheim Dr Carl van Colen Ghent University (UGhent), Ghent Belgium Dr Dorte Herzke Norwegian Institute of Air Research (NILU), Tromsø Norway Dr Kerstin Magnusson IVL Swedish Environmental Research Institute (IVL), Göteborg Sweden Prof. Geir Gabrielsen The Norwegian -

<I>Megabalanus Decorus</I>
JONES AND FOSTER: NEKTONIC EXISTENCE OF Megabalanus decorus 77 RECORDS OF NEKTONIC EXISTENCE OF THE SESSILE BARNACLE (MEGABALANUS DECORUS) M. B. JONES1 and B. A. FOSTER2 SUMMARY: The balanomorph barnacle Megabalanus decorus (Darwin, 1854) (Balanidae) is recorded attached to a plastic fish tag taken from a trevally (Caranx georgianus) collected in the Bay of Plenty (37o 53.2' S, 176o 50' E), and attached to the spine of the posterior dorsal fin of a male dogfish (Squalus acanthias L.) trawled from off the Kaikoura Peninsula (42o 25' S, 173o 42' E), New Zealand. Despite its extensive geographical distribution from the Kermadec Islands (30° S, 178o 30' E) to Macquarie Island (54o 29' S, 158o 58' E), and its usage of ships, sharks and other nektonic objects, M. decorus has not spread beyond the New Zealand region. off the Taranaki coast (B. A. Foster, pers. obs.). The INTRODUCTION present paper adds two new unusual habitat sites to Megabalanus decorus (Darwin, 1854) is a balano- the list. morph barnacle (Cirripedia, Thoracica) commonly found cast up on beaches throughout New Zealand. GEOGRAPHICAL DISTRIBUTION Its large size, compared with other New Zealand M. decorus is endemic to New Zealand waters. balanomorphs, and the rose pink colour of its shell The record of Hutton (1879) of specimens in the make it a particularly notable barnacle, and when Otago Museum from South Australia has not been attached to shells of large molluscs it can be regarded substantiated: the barnacle collection of the Otago as decorative. The morphology of this species has Museum has been very neglected, and it is not been described by Darwin (1854), Linzey (1942) and possible to identify Hutton's so-named specimens. -

Teaching About the Unintended Consequences of Plastics Tamara
From Disposable Culture to Disposable People: Teaching About the Unintended Consequences of Plastics Tamara "Sasha" Adkins, MPH Dissertation Committee Dr. Alesia Maltz, Committee Chair Core Faculty, Environmental Studies, Antioch University New England Dr. Abigail Abrash Walton, Teaching Faculty, Environmental Studies, Antioch University New England Dr. Janaki Natarajan Tschannerl, Director, SPARK Teacher Education / Educational Praxis & Director, Bapagrama Educational Center, Bangalore, India From Disposable Culture to Disposable People: Teaching About the Unintended Consequences of Plastics By Tamara "Sasha" Adkins, DISSERTATION Submitted in partial fulfillment of the requirements for the degree of Ph.D. in the Department of Environmental Studies Antioch University New England Keene, New Hampshire November 2017 Copyright 2017 Tamara “Sasha” Adkins All Rights Reserved Dedication Zendaya, To you I dedicate not only this dissertation but all my work to shape a world where no one is disposable. i Acknowledgments Alesia, my adviser and mentor for the past decade, sets a high bar for wisdom, compassion, and kindness. I will always remember you saying, "Oh, sweetie, too many sighs! If you are not having fun (with the dissertation research) then something is wrong." Thank you for not only patiently drawing out my best work, but for making the process a delightful one. Janaki, thank you for all I am learning from you in Spark and through this process. Abi joined my committee officially in my final semester, but had already been giving me encouragement and support for many years. It is much appreciated. I'd like to thank Charles Curtin, who served on my committee for a time, but due to extenuating circumstances, was not able to continue in that role. -

An Invitation to Monitor Georgia's Coastal Wetlands
An Invitation to Monitor Georgia’s Coastal Wetlands www.shellfish.uga.edu By Mary Sweeney-Reeves, Dr. Alan Power, & Ellie Covington First Printing 2003, Second Printing 2006, Copyright University of Georgia “This book was prepared by Mary Sweeney-Reeves, Dr. Alan Power, and Ellie Covington under an award from the Office of Ocean and Coastal Resource Management, National Oceanic and Atmospheric Administration. The statements, findings, conclusions, and recommendations are those of the authors and do not necessarily reflect the views of OCRM and NOAA.” 2 Acknowledgements Funding for the development of the Coastal Georgia Adopt-A-Wetland Program was provided by a NOAA Coastal Incentive Grant, awarded under the Georgia Department of Natural Resources Coastal Zone Management Program (UGA Grant # 27 31 RE 337130). The Coastal Georgia Adopt-A-Wetland Program owes much of its success to the support, experience, and contributions of the following individuals: Dr. Randal Walker, Marie Scoggins, Dodie Thompson, Edith Schmidt, John Crawford, Dr. Mare Timmons, Marcy Mitchell, Pete Schlein, Sue Finkle, Jenny Makosky, Natasha Wampler, Molly Russell, Rebecca Green, and Jeanette Henderson (University of Georgia Marine Extension Service); Courtney Power (Chatham County Savannah Metropolitan Planning Commission); Dr. Joe Richardson (Savannah State University); Dr. Chandra Franklin (Savannah State University); Dr. Dionne Hoskins (NOAA); Dr. Charles Belin (Armstrong Atlantic University); Dr. Merryl Alber (University of Georgia); (Dr. Mac Rawson (Georgia Sea Grant College Program); Harold Harbert, Kim Morris-Zarneke, and Michele Droszcz (Georgia Adopt-A-Stream); Dorset Hurley and Aimee Gaddis (Sapelo Island National Estuarine Research Reserve); Dr. Charra Sweeney-Reeves (All About Pets); Captain Judy Helmey (Miss Judy Charters); Jan Mackinnon and Jill Huntington (Georgia Department of Natural Resources).