Walls Offer Potential to Improve Urban Biodiversity
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

WIAD CONSERVATION a Handbook of Traditional Knowledge and Biodiversity
WIAD CONSERVATION A Handbook of Traditional Knowledge and Biodiversity WIAD CONSERVATION A Handbook of Traditional Knowledge and Biodiversity Table of Contents Acknowledgements ...................................................................................................................... 2 Ohu Map ...................................................................................................................................... 3 History of WIAD Conservation ...................................................................................................... 4 WIAD Legends .............................................................................................................................. 7 The Story of Julug and Tabalib ............................................................................................................... 7 Mou the Snake of A’at ........................................................................................................................... 8 The Place of Thunder ........................................................................................................................... 10 The Stone Mirror ................................................................................................................................. 11 The Weather Bird ................................................................................................................................ 12 The Story of Jelamanu Waterfall ......................................................................................................... -

Prakriti Vol 1 Issue 15
PRAKRITI, 05th October 2020, Issue 15 PRAKRITI Quest for nature Officers’ Club, CASFOS, Dehradun P R A K R I T I, 05 October, 2 0 2 0, V O L. I, I S S U E 15 FLORA OF THE WEEK Ficus virens Pakhad or Pilkhan Introduction : It is a medium sized tree which grows to a height of 23 to 28 metres in dry area and up to 30-32 metres tall in water area. In the Indian environment it has two marked growth periods: in spring (February to early May) and in monsoon rain (June to early September). It is a fig tree belonging to the group of trees known as strangler figs, because its seeds can germinate on other trees and grow to strangle and eventually kill the host tree. Morphology: Kingdom Plantae The genus Ficus falling within evergreen, semi-evergreen and deciduous species, which occupy almost any possible Order Rosales plant form, such as trees of large, medium and low growth, as well as shrubs, vines and climbers. On the Family Moraceae other hand, several species are epiphytes and other hemiepiphytes, while some are rheophytes and some other purely lithophytes. Amongst their undoubtedly distinct Genus Ficus place possess the „stranglers„ species. it initially grows on the branches of other trees from seed that have deposited frugivorous birds. From there it develops blastogenic Species virens roots that penetrate the soil. The aerial part of these roots becomes trunkish while at the same time continuing to develop blastogenic roots. Distribution: Ficus virens is found in India, Nepal, Uses: Pakistan, and South-east Asia through Malaysia, Africa, North America and Young shoots and young leaves - raw or cooked. -

Evaluation of Carrageenan Induced Anti-Inflammatory Activity of Ethanolic Extract of Bark of Ficus Virens Linn. in Swiss Albino
The Journal of Phytopharmacology 2013; 2(3): 39-43 Online at: www.phytopharmajournal.com Research Article Evaluation of Carrageenan induced anti-inflammatory ISSN 2230-480X activity of ethanolic extract of bark of Ficus virens Linn. JPHYTO 2013; 2(3): 39-43 © 2013, All rights reserved in swiss albino mice Abdul Hafeez, Dr. Upendra Jain, Pinky Sajwan, Sirish Srivastava, Amit Thakur Abdul Hafeez* Abstract Doon College of Pharmacy, Sunderpur, Saharanpur, U.P., India- The aim of the present study was to explore the probable anti-inflammatory activity of ethanolic 247001 extract of Ficus virens bark using Carrageenan induced inflammation in the mice. Swiss albino Dr. Upendra Jain mice were treated orally with normal saline (as control group) and Ficus virens extract (200 and Chandigarh College of Pharmacy, 400 mg/kg), 60 min before 0.1 mL 1% carrageenan injection. Paw volume was measured before Mohali, Punjab, India- 140110 and 1, 2, and 3 h after the injection of carrageenan. The results were expressed as the Mean ±SEM and the statistical significance of differences between groups was analyzed by One Way Pinky Sajwan Analysis of Variance (ANOVA) followed by Dunnett’s test. The sub plantar injection of Doon College of Pharmacy, Sunderpur, Saharanpur, U.P., India- carrageenan caused a time-dependent paw edema in the mice. Oral administration of Ficus 247001 virens extract (200 and 400 mg/kg) inhibited paw swelling dose-dependently at 1, 2, and 3, h after Carrageenan injection. We can conclude from the outcome of the present work that Ficus Sirish Srivastava, Amit Thakur virens extract exert an excellent anti-inflammatory effect in the mice. -

Ficus Virens Aiton, 1789
Ficus virens Aiton, 1789 Identifiants : 14123/ficvir Association du Potager de mes/nos Rêves (https://lepotager-demesreves.fr) Fiche réalisée par Patrick Le Ménahèze Dernière modification le 28/09/2021 Classification phylogénétique : Clade : Angiospermes ; Clade : Dicotylédones vraies ; Clade : Rosidées ; Clade : Fabidées ; Ordre : Rosales ; Famille : Moraceae ; Classification/taxinomie traditionnelle : Règne : Plantae ; Sous-règne : Tracheobionta ; Division : Magnoliophyta ; Classe : Magnoliopsida ; Ordre : Rosales ; Famille : Moraceae ; Genre : Ficus ; Synonymes : Ficus carolinensis Warb, Ficus cunninghamii (Miquel) Miquel, Ficus infectoria Miq, Ficus infectoria Roxb. var. lambertiana King, Ficus lacor Buch.-Ham, Ficus lucescens Blume, Urostigma lambartiana Miq, et d'autres ; Nom(s) anglais, local(aux) et/ou international(aux) : white fig, banyan, deciduous fig , Aaw, Albayi, Amchar, Biguan Rong, Chakkila, Gelong den, Hishi, Hpak-hi, Kabra, Labra, Lu huang ge shu, Neme adao, Niza bao, Nyaung-chin, Nyaung-gyin, Nyaung-shin, Pakar, Pakur, Payer, Pen-cap banyan, Phak lueat, Pilkhan, Sour buds tree, Suan bao shu ; Rapport de consommation et comestibilité/consommabilité inférée (partie(s) utilisable(s) et usage(s) alimentaire(s) correspondant(s)) : Fruit0(+x) (fruits0(+x) {chair/pulpe, peau et graines{{{(dp*)}) et feuille (jeunes(dp*) feuilles0(+x)) comestible0(+x). Détails : Les fruits sont consommés{{{0(+). La peau et les graines des fruits sont-elles bien comestibles et consommables ? (qp*). Jeunes feuilles cuites (ex. : comme potherbe) ? (qp*). Les fruits sont mangés. Les jeunes feuilles sont cuites et mangées. Ils sont également utilisés dans la soupe. Les bourgeons ou stipules sont bouillis pour donner un goût amer néant, inconnus ou indéterminés.néant, inconnus ou indéterminés. Illustration(s) (photographie(s) et/ou dessin(s)): Page 1/3 De gauche à droite : Annals of the Royal Botanic Garden, Calcutta (vol. -

How to Look at Figs
How to Look at Figs Species name: __________________________________________ Growth habit: tree or shrub or climbing vine Stems: Pith hollow or pith solid Young twigs: hairy or hairless Roots: aerial roots present buttress roots present Leaves Leaf blade shape: or draw shape: Leaf margin: lobed or entire (smooth margins) Leaf tips: long and tapering (acute, with a drip tip) or blunt (obtuse) Leaf base: tapering to petiole or truncate symmetrical or asymmetrical Stipules: stipule or terminal bud length: ________ stipules fused into 1 or two stipules per node Leaf venation: 1 main vein or several veins from the base side veins few or side veins many Venation angle: parallel or acute or obtuse Leaf hairs: leaves hairless (glabrous) or covered with fne hairs Leaf hair color: golden white yellow other: ________ Figs cauliforous fgs present on woody stems or found in axils of leaves fgs single or paired sessile or stalked Fig diameter:____________ Fig color:____________ Fig hairs: leaves hairless (glabrous) or covered with fne hairs Fig hair color: golden white yellow other: ________ Fig surface: smooth hairy (pubescent) ribbed fecks of white bracts present on body or base of fgs Fig internal surface: size:____________ color: ____________ Other Notable Features: Characteristics and Taxonomy of Ficus Family: Moraceae Genus Ficus = ~850 species Original publication: Linnaeus, Species Plantarum 2: 1059. 1753 Subgenera and examples of commonly grown species: Subgenus Ficus (~350 species) Africa, Asia, Australasia, Mediterranean Mostly gynodioecious, no bracts among the fowers, free standing trees and shrubs, cauliforous or not, pollination passive Ficus carica L. – common fg Ficus deltoidea Jack – mistletoe fg Subgenus Sycidium (~100 species) Africa, Asia, Australasia Mostly gynodioecious, no bracts among the fowers, free standing trees and shrubs, cauliforous or not, pollination passive Ficus coronata Spin - creek sandpaper fg Ficus fraseri Miq. -

Boigu Island in the Torres Strait
PROFILE FOR MANAGEMENT OF THE HABITATS AND RELATED ECOLOGICAL AND CULTURAL RESOURCE VALUES OF IAMA ISLAND January 2013 Prepared by 3D Environmental for Torres Strait Regional Authority Land & Sea Management Unit Cover image: 3D Environmental (2013) EXECUTIVE SUMMARY Iama Island, which occupies a total area of 186 ha, is formed on a pile of granitic basement rocks that outcrop to a height of 68m. The rocky interior of the island is fringed by a number of younger landform features including beach ridges and broad flats formed from estuarine mud. A total of 14 vegetation communities, within 6 intact broad vegetation groups and 11 regional ecosystems are recognised across the island, representing approximately 14% of regional ecosystems recorded across the broader Torres Strait Island landscape. The most widespread vegetation type on the island other than mangrove is an open forest formation dominated by earlobe wattle (Acacia auriculiformis) (Thulup in local dialect). This is a unique habitat type found only on Iama. There are currently 259 flora species recorded on the island which comprises 202 (79%) native taxa, with 57 (21%) introduced species. This represents approximately 20% of the known flora for the Torres Strait Island group. One species of palm Arenga australasica, is listed as Vulnerable under Federal and State legislation with three species Dolichandrone spathacea (a shrub on mangrove margins), Operculina brownii (a vine of vine thickets) and Neololeba atra (a bamboo on vine thicket margins), listed as Near-Threatened under the Queensland Nature Conservation Act 1992. A further 12 species are considered to have significance at a local and regional level. -
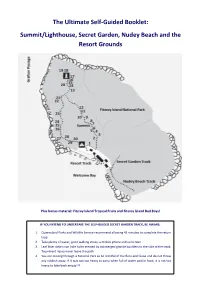
The Ultimate Self-Guided Booklet: Summit/Lighthouse, Secret Garden, Nudey Beach and the Resort Grounds
The Ultimate Self-Guided Booklet: Summit/Lighthouse, Secret Garden, Nudey Beach and the Resort Grounds Plus bonus material: Fitzroy Island Tropical Fruits and Fitzroy Island Bad Boys! IF YOU INTEND TO UNDERTAKE THE SELF-GUIDED SECRET GARDEN TRACK, BE AWARE: 1. Queensland Parks and Wildlife Service recommend allowing 45 minutes to complete the return loop 2. Take plenty of water, good walking shoes, a mobile phone and sunscreen 3. Leaf litter debris can hide holes created by submerged granite boulders to the side of the track. To prevent injury never leave the path 4. You are moving through a National Park so be mindful of the flora and fauna and do not throw any rubbish away. If it was not too heavy to carry when full of water and/or food, it is not too heavy to take back empty!!! The information in this booklet is dedicated to all those that have called Fitzroy Island home, from the island’s original inhabitants, the Gunggandji people, to the military and lightkeepers who each played their role in shaping the island’s fate. The Gunggandji were walking these paths before paths existed, living off these trees and surviving in this forest for thousands of years before the first European explorers ever set foot on this land. When war threatened our shores the men of the No. 28 Radar Station stepped up to protect these lands; their influence can still be seen across the island today as can the Lightkeepers that came after them. Each served their country in isolation in lieu of modern comforts. -

Composition of Trees Grown Surrounding Water Springs at Two Areas in Purwosari Pasuruan, East Java
THE JOURNAL OF TROPICAL LIFE SCIENCE OPEN ACCESS Freely available online VOL. 2, NO. 2, pp. 110 – 118, September, 2012 Composition of Trees Grown Surrounding Water Springs at Two Areas in Purwosari Pasuruan, East Java Soejono Purwodadi Botanic Garden-Indonesian Institute of Sciences ABSTRACT The aim of the research was to find out the composition of trees grown surrounding water springs at two areas in Purwosari, Pasuruan, East Java. Eleven plots for each area were observed. The data were analyzed using Mueller-Dombois’s method to calculate their importance value indexes, while Shannon-Wiener’s formula was used for determining the diversity index. The coordinate and altitude of every water spring or its group’s site was determined using Geographical Position System (GPS) to know their positions on the map. The result indicated that there were at least 30 families, 49 genera which consisted of 68 species of trees grown surrounding water springs at the first area with 5.49 of diversity index, while the second area, consisted of 34 families, 63 genera and 79 species of trees with 5.24 diversity index. The diversity of trees species from Moraceae was the highest among other families, both at the first and the second area, whereas, trees species having a significant important value index included Bambusa blumeana, Dendrocalamus asper, Ficus racemosa, Horsfieldia irya and Ficus virens. The position of the springs in the two areas within the sub-districts of Purwosari is in the range of 7º44'448 " south latitude; 112º44'353" east longitude up to 7º46'339 " south latitude; 112º41’190" east longitude at an altitude between 251 and 522 m above sea level. -

Furnieles Nuñez Hector Javier.Pdf
ESTUDIO QUIMIOTAXONÓMICO Y EVALUACIÓN DE LA ACTIVIDAD ANTIOXIDANTE DE EXTRACTOS ETANÓLICOS FOLIARES DE CUATRO ESPECIES DEL GÉNERO Ficus L. (Moraceae), PLANETA RICA (CÓRDOBA- COLOMBIA). HÉCTOR JAVIER FURNIELES NÚÑEZ UNIVERSIDAD DE CÓRDOBA FACULTAD DE CIENCIAS BÁSICAS PROGRAMA DE BIOLOGÍA 2020 ESTUDIO QUIMIOTAXONÓMICO Y EVALUACIÓN DE LA ACTIVIDAD ANTIOXIDANTE DE EXTRACTOS ETANÓLICOS FOLIARES DE CUATRO ESPECIES DEL GÉNERO Ficus L. (Moraceae), PLANETA RICA (CÓRDOBA- COLOMBIA). HÉCTOR JAVIER FURNIELES NÚÑEZ Investigador Principal DIRECTORES JORGE ENRIQUE ARIAS, M.Sc. Docente programa de Biología Universidad de Córdoba MARY CECILIA MONTAÑO, PhD. Docente programa de Química Universidad de Córdoba UNIVERSIDAD DE CÓRDOBA FACULTAD DE CIENCIAS BÁSICAS PROGRAMA DE BIOLOGÍA 2020 AGRADECIMIENTOS Agradezco principalmente a Dios, por brindarme sabiduría, fuerza y constancia para lograr culminar con éxito este importante objetivo en mi vida. A mi familia por su apoyo y confianza, a la tía universal Nadys Núñez Ortega, gracias por tanto. A mis queridos padres Héctor Antonio Furnieles y Nerys Núñez Ortega, a mis hermanas Heidy Furnieles Núñez y Margarita Furnieles Núñez, por el apoyo incondicional y acompañamiento constante, aun en los momentos difíciles de esta hermosa travesía investigativa. A la Doctora Mary Montaño y al Magister Jorge Arias, por asumir la dirección de este proyecto, infinitamente agradecido por sus orientaciones, por compartir sus importantes y valiosos conocimientos científicos, que permitieron en mí el crecimiento profesional y científico con pensamiento crítico en las ciencias exactas. A los jurados, Doctora Rosalba Ruiz Vega y Magister Emmy Luz Sánchez, por sus valiosas recomendaciones. A mis compañeros de estudio, mil gracias por su amistad, hermandad y compañerismo. Gracias por animarme a seguir en las dificultades. -

Andaman & Nicobar Islands, India
RESEARCH Vol. 21, Issue 68, 2020 RESEARCH ARTICLE ISSN 2319–5746 EISSN 2319–5754 Species Floristic Diversity and Analysis of South Andaman Islands (South Andaman District), Andaman & Nicobar Islands, India Mudavath Chennakesavulu Naik1, Lal Ji Singh1, Ganeshaiah KN2 1Botanical Survey of India, Andaman & Nicobar Regional Centre, Port Blair-744102, Andaman & Nicobar Islands, India 2Dept of Forestry and Environmental Sciences, School of Ecology and Conservation, G.K.V.K, UASB, Bangalore-560065, India Corresponding author: Botanical Survey of India, Andaman & Nicobar Regional Centre, Port Blair-744102, Andaman & Nicobar Islands, India Email: [email protected] Article History Received: 01 October 2020 Accepted: 17 November 2020 Published: November 2020 Citation Mudavath Chennakesavulu Naik, Lal Ji Singh, Ganeshaiah KN. Floristic Diversity and Analysis of South Andaman Islands (South Andaman District), Andaman & Nicobar Islands, India. Species, 2020, 21(68), 343-409 Publication License This work is licensed under a Creative Commons Attribution 4.0 International License. General Note Article is recommended to print as color digital version in recycled paper. ABSTRACT After 7 years of intensive explorations during 2013-2020 in South Andaman Islands, we recorded a total of 1376 wild and naturalized vascular plant taxa representing 1364 species belonging to 701 genera and 153 families, of which 95% of the taxa are based on primary collections. Of the 319 endemic species of Andaman and Nicobar Islands, 111 species are located in South Andaman Islands and 35 of them strict endemics to this region. 343 Page Key words: Vascular Plant Diversity, Floristic Analysis, Endemcity. © 2020 Discovery Publication. All Rights Reserved. www.discoveryjournals.org OPEN ACCESS RESEARCH ARTICLE 1. -

Ficus Religiosa Linn.(Peepal): a Phyto-Pharmacological
INTERNATIONAL JOURNAL OF PHARMACEUTICAL AND CHEMICAL SCIENCES Review Article Ficus religiosa Linn. (Peepal): A Phyto-Pharmacological Review Sunita Panchawat Department of Pharmacognosy, Bhupal Noble’s Girls’ College of Pharmacy, Udaipur, Rajasthan, India. __________________________________________________________________ ABSTRACT Ficus religiosa is known to be a native Indian tree. It is a familiar sight in Hindu temples, Buddhist monasteries and shrines, villages and at roadsides. It is known to be a sacred plant in India and since ancient times it is widely being used to treat various ailments like skin diseases, heart diseases, constipation, dysentery, snakebite and important constituent of various traditional herbal preparations like shankha vati, chandraprabha vati, kaminivindravan rasa. In recent studies Ficus religiosa has been investigated for presence of a wide range of phytoconstituents viz. glycosides, flavonoids, tannin, terpenoids and sterol. This compilation provides comprehensive review of its ethnopharmacological use, phytochemistry, phytopathology, pharmacological activities, traditional and medicinal use. Keywords: Ficus religiosa, phytochemistry, Phytopathology, antioxidant, pharmacological activities. INTRODUCTION Ficus religiosa is a variety of fig tree that Vedic history was already known as the bodhi tree, The Brahma Purana and the Padma even before Gautama Buddha sat under Purana, relate how once, when the its branches meditating and achieved demons defeated the Gods, Vishnu hide enlightenment. It is the oldest depicted in the peepal. Therefore spontaneous tree in Indian art and literature and it can worship to Vishnu can be offered to a be said that this is the mythical 'World peepal without needing his image or Tree' or the 'Tree of Life' of the Indian subcontinent. This plant is considered temple. The Skanda Purana also sacred by the followers of Hinduism, considers the peepal, a symbol of Vishnu. -

Apg-Iii 2009
APG-III 2009 18S rDNA rbcL atpB atp1 matR + 61 genes de 45 táxons 75% EUDICOTILEDÔNEAS: 41 ordens Amborellales Nymphaeales APG III 2009 Austrobaileyales Chloranthales Magnoliídeas Monocotiledôneas Ceratophyllales Eudicotiledôneas (ou Tricolpadas) pólen tricolpado Hamamelis pólen Euphorbia tricolporado EUDICOTILEDÔNEAS Aesculus Cobaea Silene Cucurbita pólen porado Cucumis APG-III 2009 18S rDNA rbcL atpB atp1 matR + 61 genes APG-III 2009 Soltis et al. 2009 Ranunculales Proteales 2-meria Trochodendrales Gunnerales ROSÍDEAS 5-meria Santalales Caryophyllales Eudicots nucleares ASTERÍDEAS PENTAPETALE Ranunculales Proteales 2-meria Trochodendrales Gunnerales ROSÍDEAS Santalales Acanthaceae 4-5-meria Caryophyllales ASTERÍDEAS Aesculus Acanthaceae Euphorbia pólen colporado a porado Ranunculales Eudicotiledôneas Proteales basais Trochodendrales Gunnerales ROSÍDEAS c.140 famílias c. 68.000 spp. (1/4 das angiospermas) 4-5-meria Santalales pólen Caryophyllales colporado ASTERÍDEAS Eudicots nucleares c. 96 famílias ROSÍDEAS - sinapomorfias estípulas redução do endosperma diplostemonia APG-III ROSÍDEAS 2009 18S rDNA rbcL atpB fabídeas atp1 matR estípulas malvídeas redução do endosperma redução ROSÍDEAS FABÍDEAS 8 ordens/ c. 76 famílias Sinapomorfias macromoleculares! Clado fixador de N2 (Frankia ou Rhizobium) Stevens 2015 10 famílias ROSALES 6300 spp. hipanto CANNABACEAE ROSACEAE ROSACEAE Prunus ROSALES 1 carpelo Prunus ROSACEAE ROSALES muitos carpelos Fragaria ROSACEAE Rosa rugosa Rosa Malus robusta ROSACEAE ROSALES ROSACEAE Malus robusta ROSALES Flor