Smallest Unit of Life: Cell Biology 2 Isabella Ellinger and Adolf Ellinger
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Glossary - Cellbiology
1 Glossary - Cellbiology Blotting: (Blot Analysis) Widely used biochemical technique for detecting the presence of specific macromolecules (proteins, mRNAs, or DNA sequences) in a mixture. A sample first is separated on an agarose or polyacrylamide gel usually under denaturing conditions; the separated components are transferred (blotting) to a nitrocellulose sheet, which is exposed to a radiolabeled molecule that specifically binds to the macromolecule of interest, and then subjected to autoradiography. Northern B.: mRNAs are detected with a complementary DNA; Southern B.: DNA restriction fragments are detected with complementary nucleotide sequences; Western B.: Proteins are detected by specific antibodies. Cell: The fundamental unit of living organisms. Cells are bounded by a lipid-containing plasma membrane, containing the central nucleus, and the cytoplasm. Cells are generally capable of independent reproduction. More complex cells like Eukaryotes have various compartments (organelles) where special tasks essential for the survival of the cell take place. Cytoplasm: Viscous contents of a cell that are contained within the plasma membrane but, in eukaryotic cells, outside the nucleus. The part of the cytoplasm not contained in any organelle is called the Cytosol. Cytoskeleton: (Gk. ) Three dimensional network of fibrous elements, allowing precisely regulated movements of cell parts, transport organelles, and help to maintain a cell’s shape. • Actin filament: (Microfilaments) Ubiquitous eukaryotic cytoskeletal proteins (one end is attached to the cell-cortex) of two “twisted“ actin monomers; are important in the structural support and movement of cells. Each actin filament (F-actin) consists of two strands of globular subunits (G-Actin) wrapped around each other to form a polarized unit (high ionic cytoplasm lead to the formation of AF, whereas low ion-concentration disassembles AF). -

Bacterial Cell Membrane
BACTERIAL CELL MEMBRANE Dr. Rakesh Sharda Department of Veterinary Microbiology NDVSU College of Veterinary Sc. & A.H., MHOW CYTOPLASMIC MEMBRANE ➢The cytoplasmic membrane, also called a cell membrane or plasma membrane, is about 7 nanometers (nm; 1/1,000,000,000 m) thick. ➢It lies internal to the cell wall and encloses the cytoplasm of the bacterium. ➢It is the most dynamic structure of a prokaryotic cell. Structure of cell membrane ➢The structure of bacterial plasma membrane is that of unit membrane, i.e., a fluid phospholipid bilayer, composed of phospholipids (40%) and peripheral and integral proteins (60%) molecules. ➢The phospholipids of bacterial cell membranes do not contain sterols as in eukaryotes, but instead consist of saturated or monounsaturated fatty acids (rarely, polyunsaturated fatty acids). ➢Many bacteria contain sterol-like molecules called hopanoids. ➢The hopanoids most likely stabilize the bacterial cytoplasmic membrane. ➢The phospholipids are amphoteric molecules with a polar hydrophilic glycerol "head" attached via an ester bond to two non-polar hydrophobic fatty acid tails. ➢The phospholipid bilayer is arranged such that the polar ends of the molecules form the outermost and innermost surface of the membrane while the non-polar ends form the center of the membrane Fluid mosaic model ➢The plasma membrane contains proteins, sugars, and other lipids in addition to the phospholipids. ➢The model that describes the arrangement of these substances in lipid bilayer is called the fluid mosaic model ➢Dispersed within the bilayer are various structural and enzymatic proteins, which carry out most membrane functions. ➢Some membrane proteins are located and function on one side or another of the membrane (peripheral proteins). -

Intelligent Design, Abiogenesis, and Learning from History: Dennis R
Author Exchange Intelligent Design, Abiogenesis, and Learning from History: Dennis R. Venema A Reply to Meyer Dennis R. Venema Weizsäcker’s book The World View of Physics is still keeping me very busy. It has again brought home to me quite clearly how wrong it is to use God as a stop-gap for the incompleteness of our knowledge. If in fact the frontiers of knowledge are being pushed back (and that is bound to be the case), then God is being pushed back with them, and is therefore continually in retreat. We are to find God in what we know, not in what we don’t know; God wants us to realize his presence, not in unsolved problems but in those that are solved. Dietrich Bonhoeffer1 am thankful for this opportunity to nature, is the result of intelligence. More- reply to Stephen Meyer’s criticisms over, this assertion is proffered as the I 2 of my review of his book Signature logical basis for inferring design for the in the Cell (hereafter Signature). Meyer’s origin of biological information: if infor- critiques of my review fall into two gen- mation only ever arises from intelli- eral categories. First, he claims I mistook gence, then the mere presence of Signature for an argument against bio- information demonstrates design. A few logical evolution, rendering several of examples from Signature make the point my arguments superfluous. Secondly, easily: Meyer asserts that I have failed to refute … historical scientists can show that his thesis by not providing a “causally a presently acting cause must have adequate alternative explanation” for the been present in the past because the origin of life in that the few relevant cri- proposed candidate is the only known tiques I do provide are “deeply flawed.” cause of the effect in question. -

The Endomembrane System and Proteins
Chapter 4 | Cell Structure 121 Endosymbiosis We have mentioned that both mitochondria and chloroplasts contain DNA and ribosomes. Have you wondered why? Strong evidence points to endosymbiosis as the explanation. Symbiosis is a relationship in which organisms from two separate species depend on each other for their survival. Endosymbiosis (endo- = “within”) is a mutually beneficial relationship in which one organism lives inside the other. Endosymbiotic relationships abound in nature. We have already mentioned that microbes that produce vitamin K live inside the human gut. This relationship is beneficial for us because we are unable to synthesize vitamin K. It is also beneficial for the microbes because they are protected from other organisms and from drying out, and they receive abundant food from the environment of the large intestine. Scientists have long noticed that bacteria, mitochondria, and chloroplasts are similar in size. We also know that bacteria have DNA and ribosomes, just like mitochondria and chloroplasts. Scientists believe that host cells and bacteria formed an endosymbiotic relationship when the host cells ingested both aerobic and autotrophic bacteria (cyanobacteria) but did not destroy them. Through many millions of years of evolution, these ingested bacteria became more specialized in their functions, with the aerobic bacteria becoming mitochondria and the autotrophic bacteria becoming chloroplasts. The Central Vacuole Previously, we mentioned vacuoles as essential components of plant cells. If you look at Figure 4.8b, you will see that plant cells each have a large central vacuole that occupies most of the cell's area. The central vacuole plays a key role in regulating the cell’s concentration of water in changing environmental conditions. -

The Hidden Kingdom
INTRODUCTION Fungi—The Hidden Kingdom OBJECTIVE • To provide students with basic knowledge about fungi Activity 0.1 BACKGROUND INFORMATION The following text provides an introduction to the fungi. It is written with the intention of sparking curiosity about this GRADES fascinating biological kingdom. 4-6 with a K-3 adaptation TEACHER INSTRUCTIONS TYPE OF ACTIVITY 1. With your class, brainstorm everything you know about fungi. Teacher read/comprehension 2. For younger students, hand out the question sheet before you begin the teacher read and have them follow along and MATERIALS answer the questions as you read. • copies of page 11 3. For older students, inform them that they will be given a • pencils brainteaser quiz (that is not for evaluation) after you finish reading the text. VOCABULARY 4. The class can work on the questions with partners or in groups bioremediation and then go over the answers as a class. Discuss any chitin particularly interesting facts and encourage further fungi independent research. habitat hyphae K-3 ADAPTATION kingdom 1. To introduce younger students to fungi, you can make a KWL lichens chart either as a class or individually. A KWL chart is divided moulds into three parts. The first tells what a student KNOWS (K) mushrooms about a subject before it is studied in class. The second part mycelium tells what the student WANTS (W) to know about that subject. mycorrhizas The third part tells what the child LEARNED (L) after studying nematodes that subject. parasitic fungi 2. Share some of the fascinating fungal facts presented in the photosynthesis “Fungi—The Hidden Kingdom” text with your students. -

Framing Major Prebiotic Transitions As Stages of Protocell Development: Three Challenges for Origins-Of-Life Research
Framing major prebiotic transitions as stages of protocell development: three challenges for origins-of-life research Ben Shirt-Ediss1, Sara Murillo-Sánchez2,3 and Kepa Ruiz-Mirazo*2,3 Commentary Open Access Address: Beilstein J. Org. Chem. 2017, 13, 1388–1395. 1Interdisciplinary Computing and Complex BioSystems Group, doi:10.3762/bjoc.13.135 University of Newcastle, UK, 2Dept. Logic and Philosophy of Science, University of the Basque Country, Spain and 3Biofisika Institute Received: 16 February 2017 (CSIC, UPV-EHU), Spain Accepted: 27 June 2017 Published: 13 July 2017 Email: Kepa Ruiz-Mirazo* - [email protected] This article is part of the Thematic Series "From prebiotic chemistry to molecular evolution". * Corresponding author Guest Editor: L. Cronin Keywords: functional integration; origins of life; prebiotic evolution; protocells © 2017 Shirt-Ediss et al.; licensee Beilstein-Institut. License and terms: see end of document. Abstract Conceiving the process of biogenesis as the evolutionary development of highly dynamic and integrated protocell populations provides the most appropriate framework to address the difficult problem of how prebiotic chemistry bridged the gap to full-fledged living organisms on the early Earth. In this contribution we briefly discuss the implications of taking dynamic, functionally inte- grated protocell systems (rather than complex reaction networks in bulk solution, sets of artificially evolvable replicating molecules, or even these same replicating molecules encapsulated in passive compartments) -

Standard 2: CELL BIOLOGY – REVIEW of BASICS
Standard 2: CELL BIOLOGY – REVIEW OF BASICS CELL PART OR TYPE OF CELL WHERE FOUND WHAT DOES IT FUNCTION: MISCELLANEOUS ORGANELLE Prokaryotic cell Plant cell LOOK LIKE: Job it does in INFORMATION: things Eukaryotic cell Animal cell Describe or Draw the cell such as color, what it is Both Both made of, size, etc. plasma/cell See diagram Holds cell together Phospholipid bilayer with membrane both both Regulates what goes proteins in/out of cell Semipermeable cytoplasm both Clear thick jelly- Supports/protects both like material in cell cell organelles See diagram Control center nucleus eukaryotic both Contains DNA See diagram Where proteins are ribosome both both made See diagram Process proteins Golgi complex eukaryotic both that go to other /apparatus parts of cell Membrane-bound Digests materials lysosome eukaryotic animal sac of digestive within the cell enzymes Membrane-bound Stores water, food, One large one in plants vacuole eukaryotic both storage area waste and dissolved Many smaller ones in minerals animals endoplasmic Network of Transport materials Can be rough (with reticulum eukaryotic both membrane tubes throughout the cell ribosomes attached) or smooth (without ribosomes) See diagram Where cell respiration Called Powerhouse of cell mitochondria eukaryotic both occurs (releases Makes ATP from energy for cell to use) breaking down glucose See diagram Where photosynthesis Contains chlorophyll chloroplast eukaryotic plant takes place Converts light energy into chemical energy in glucose Some pro- and plant (also fungi Rigid structure -

Introduction to the Cell Cell History Cell Structures and Functions
Introduction to the cell cell history cell structures and functions CK-12 Foundation December 16, 2009 CK-12 Foundation is a non-profit organization with a mission to reduce the cost of textbook materials for the K-12 market both in the U.S. and worldwide. Using an open-content, web-based collaborative model termed the “FlexBook,” CK-12 intends to pioneer the generation and distribution of high quality educational content that will serve both as core text as well as provide an adaptive environment for learning. Copyright ©2009 CK-12 Foundation This work is licensed under the Creative Commons Attribution-Share Alike 3.0 United States License. To view a copy of this license, visit http://creativecommons.org/licenses/by-sa/3.0/us/ or send a letter to Creative Commons, 171 Second Street, Suite 300, San Francisco, California, 94105, USA. Contents 1 Cell structure and function dec 16 5 1.1 Lesson 3.1: Introduction to Cells .................................. 5 3 www.ck12.org www.ck12.org 4 Chapter 1 Cell structure and function dec 16 1.1 Lesson 3.1: Introduction to Cells Lesson Objectives • Identify the scientists that first observed cells. • Outline the importance of microscopes in the discovery of cells. • Summarize what the cell theory proposes. • Identify the limitations on cell size. • Identify the four parts common to all cells. • Compare prokaryotic and eukaryotic cells. Introduction Knowing the make up of cells and how cells work is necessary to all of the biological sciences. Learning about the similarities and differences between cell types is particularly important to the fields of cell biology and molecular biology. -

BIOSC 0805: the HUMAN BODY Department of Biological Sciences University of Pittsburgh
Syllabus: Biosc 0805, The Human Body BIOSC 0805: THE HUMAN BODY Department of Biological Sciences University of Pittsburgh Faculty Zuzana Swigonova, Ph.D. Office: 356 Langley Hall (Third floor, the bridge between Clapp and Langley halls) tel.: 412-624-3288; email: [email protected] Office hours Office hours: Mondays 10:00 – 11:30 AM, 356 Langley Hall Wednesdays 1:00 – 2:30 PM, 356 Langley Hall Office hours by appointment can be arranged by email. Lecture Time Tuesdays & Thursdays, 2:30 – 3:45 PM, 169 Crawford Hall Course objectives This is a course in human biology and physiology for students not majoring in biology. The goal is to provide students with an understanding of fundamental principles of life with an emphasis on the human body. We will start by covering basic biochemistry and cell biology and then move on to the structure and function of human organ systems. An essential part of the course is a discussion of health issues of general interest, such as infectious, autoimmune and neurodegenerative diseases; asthma and allergy; nutrition and health; stem cells research and cloning; and methods of contraception and reproductive technologies. Textbook • Biology. A guide to the natural world, by David Krogh. Pearson, Benjamin Cummings Publishing Company. (ISBN#:0-558-65495-9). This is custom made textbook that includes only the parts of the original edition that are covered in the course. It is available in the Pitt bookstore. • You can also use the full 4th or 3rd edition, however, be aware that the chapters in earlier editions are rearranged in a different order and may be lacking some parts included in the later edition. -

MCB5068 Intro
Goals For MCB 5068 1. Obtain a solid foundation of knowledge in cell biology 2. Obtain a working knowledge of available techniques. 3. Be able to critically read and evaluate the scientific literature. 4. Be able to define and investigate a biological problem. Molecular Cell Biology 5068 To Do: Visit website: www.mcb5068.wustl.edu Sign up for course. Check out Self Assessment homework under Mercer- Introduction Visit Discussion Sections: Read “Official” Instructions TA’s: 1st Session: Nana Owusu-Boaitey [email protected] 2nd Session: Shankar Parajuli [email protected] 3rd Session: Jeff Kremer [email protected] What is Cell Biology? biochemistry genetics cytology physiology Molecular Cell Biology CELL BIOLOGY/MICROSCOPE Microscope first built in 1595 by Hans and Zacharias Jensen in Holland Zacharias Jensen CELL BIOLOGY/MICROSCOPE Robert Hooke accomplished in physics, astronomy, chemistry, biology, geology, and architecture. Invented universal joint, iris diaphragm, anchor escapement & balance spring, devised equation describing elasticity (“Hooke’s Law”). In 1665 publishes Micrographia CELL BIOLOGY/MICROSCOPE Robert Hooke . “I could exceedingly plainly perceive it to be all perforated and porous, much like a Honey- comb, but that the pores of it were not regular. these pores, or cells, . were indeed the first microscopical pores I ever saw, and perhaps, that were ever seen, for I had not met with any Writer or Person, that had made any mention of them before this. .” CELL BIOLOGY/MICROSCOPE Antony van Leeuwenhoek (1632-1723) CELL BIOLOGY/MICROSCOPE Antony van Leeuwenhoek (1632-1723) a tradesman of Delft, Holland, in 1673, with no formal training, makes some of the most important discoveries in biology. -
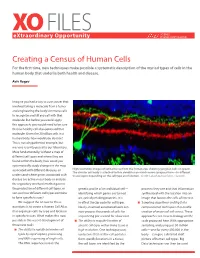
Creating a Census of Human Cells
XO FILES SCIENCE eXtraordinary Opportunity PHILANTHROPY ALLIANCE Creating a Census of Human Cells For the first time, new techniques make possible a systematic description of the myriad types of cells in the human body that underlie both health and disease. Aviv Regev Imagine you had a way to cure cancer that involved taking a molecule from a tumor and engineering the body’s immune cells to recognize and kill any cell with that molecule. But before you could apply this approach, you would need to be sure that no healthy cell also expressed that molecule. Given the 20 trillion cells in a human body, how would you do that? This is not a hypothetical example, but was one recently posed to our laboratory. More fundamentally, without a map of different cell types and where they are found within the body, how could you systematically study changes in the map associated with different diseases, or High resolution images of retinal tissue from the human eye, showing ganglion cells (in green). The circular cell body is attached to thin dendrites on which nerve synapses form—in different understand where genes associated with tissue layers depending on the cell type and function. Credit: Lab of Joshua Sanes, Harvard. disease are active in our body or analyze the regulatory mechanisms that govern the production of different cell types, or genetic profile of an individual cell— proteins they use and that information sort out how different cell types combine identifying which genes are turned synthesized with the location into an to form specific tissues? on, actively making proteins. -

Chapter 32 FOREIGN BODIES of the HEAD, NECK, and SKULL BASE
Foreign Bodies of the Head, Neck, and Skull Base Chapter 32 FOREIGN BODIES OF THE HEAD, NECK, AND SKULL BASE RICHARD J. BARNETT, MD* INTRODUCTION PENETRATING NECK TRAUMA Anatomy Emergency Management Clinical Examination Investigations OPERATIVE VERSUS NONOPERATIVE MANAGEMENT Factors in the Deployed Setting Operative Management Postoperative Care PEDIATRIC INJURIES ORBITAL FOREIGN BODIES SUMMARY CASE PRESENTATIONS Case Study 32-1 Case Study 32-2 Case Study 32-3 Case Study 32-4 Case Study 32-5 Case Study 32-6 *Lieutenant Colonel, Medical Corps, US Air Force; Chief of Facial Plastic Surgery/Otolaryngology, Eglin Air Force Base Department of ENT, 307 Boatner Road, Suite 114, Eglin Air Force Base, Florida 32542-9998 423 Otolaryngology/Head and Neck Combat Casualty Care INTRODUCTION The mechanism and extent of war injuries are sig- other military conflicts. In a study done in Croatia with nificantly different from civilian trauma. Many of the 117 patients who sustained penetrating neck injuries, wounds encountered are unique and not experienced about a quarter of the wounds were from gunshots even at Role 1 trauma centers throughout the United while the rest were from shell or bomb shrapnel.1 The States. Deployed head and neck surgeons must be injury patterns resulting from these mechanisms can skilled at performing an array of evaluations and op- vary widely, and treating each injury requires thought- erations that in many cases they have not performed in ful planning to achieve a successful outcome. a prior setting. During a 6-month tour in Afghanistan, This chapter will address penetrating neck injuries all subspecialties of otolaryngology were encountered: in general, followed specifically by foreign body inju- head and neck (15%), facial plastic/reconstructive ries of the head, face, neck, and skull base.