Otieno Et Al 2015 Accepted Manuscript
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Insect Fauna Associated with Pigeon Pea (Cajanus Cajan
Journal of Pharmacognosy and Phytochemistry 2019; 8(4): 766-768 E-ISSN: 2278-4136 P-ISSN: 2349-8234 JPP 2019; 8(4): 766-768 Insect fauna associated with pigeon pea (Cajanus Received: 01-05-2019 Accepted: 04-06-2019 cajan L.) in Odisha D Padhy Department of Entomology, D Padhy, CR Satapathy and RN Mohapatra MSSSOA, Paralakhemundi, Centurion University of Abstract Technology and Management, Field experiments were conducted during Kharif, 2017-2018 undertaken at Entomology Research Station Odisha, India of Orissa University of Agriculture and Technology, Bhubaneswar to study the occurrence of different insect pollinators on Pigeon pea (Cajanus cajan L.). Though Pigeon pea is a self pollinated crop with CR Satapathy cleistogamous type of flower, it is visited by an array of insects. Under agroclimatic conditions of AICRP on Honeybees and Pollinators, Department of Odisha, It attracted eleven species of important pollinators represented by family Apidae (45.5%), Entomology, College of Megachilidae (36.4%) and Vespidae (18.2%) all belonging to a single order hymenoptera barring stay Agriculture, Bhubaneswar, population of Dipterans and Lepidopterans. Species wise diversity indicated that Megachile lanata Fab. Odisha University of Agriculture was the most dominant one (29.1%) followed by Megachile disjuncta Fab. (20.5%), Tetragonula and Technology, Odisha, India iridipennis Smith (14.2%), Apis cerana indica Fab. (10.95%) Megachile bicolor Fab. (10.7%), Xylocopa latipes Drury (8.5%), Megachile hera Bingham (2.1%), Eumenes spp. (1.7%), Xylocopa aestuans Lin. RN Mohapatra (1.2%), Vespa tropica Lin. (0.8%) and Apis dorsata Fab. (0.2%). It is inferred from the present AICRP on Honeybees and investigation that Pigeon pea is an ideal crop supporting an array of pollinators, most important being the Pollinators, Department of M. -

Hymenoptera) Associated with Crops and Ornamental Plant in Obafemi Awolowo University, Nigeria
International Journal of Zoology and Applied Biosciences ISSN: 2455-9571 Volume 4, Issue 5, pp: 195-206, 2019 http://www.ijzab.com https://doi.org/10.5281/zenodo Research Article DIVERSITY ASSESSMENT OF BEES (HYMENOPTERA) ASSOCIATED WITH CROPS AND ORNAMENTAL PLANT IN OBAFEMI AWOLOWO UNIVERSITY, NIGERIA Oyewole O.A.1, Oyelade O.J.1* and Ogbogu S.S.2 1Natural History Museum, Obafemi Awolowo University, Ile-Ife, Nigeria, 2Department of Zoology, Obafemi Awolowo University, Ile-Ife, Nigeria Article History: Received 16th May 2019; Accepted 27th June 2019; Published 9th October 2019 ABSTRACT This study investigated the various species of bees in the Order Hymenoptera responsible for pollinating crop and ornamental plants in Obafemi Awolowo University (O.A.U.), Ile-Ife, and the study covered habitat preference, abundance and distribution of the bee species in various seasons. This was with a view to identifying and documenting the diversity of bees pollinating agricultural and ornamental plants for a meaningful conservation and management of insects in Nigeria. Bees collection was done using standard sweep net, to collect free range bees species on the sites. Standard yellow pan traps were also set using pineapple as bait to entice bees. Trapped bees were collected and baits were changed on daily basis. The bees were described and identified to species level using existing bee taxonomic keys. Host plants visited by the bee species were identified in the Natural History Museum Herbarium (UNIFEM) and Department of Botany Herbarium (IFE) in Obafemi Awolowo University, Ile-Ife. Paleontological Statistic Software Package (PAST) was used to analyze the collected data. -
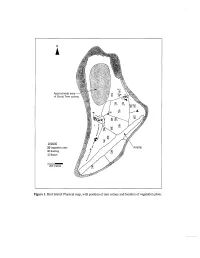
Figure 1. Bird Island: Physical Map, with Position of Tern Colony and Location of Vegetation Plots
Approximate of Sooty Ter~ Leqend (XI Vegetat~onplots 9 Building 6 Beach 200 metres Figure 1. Bird Island: Physical map, with position of tern colony and location of vegetation plots. BIRD MICHAEL J. HILL', TERENCE M. VEL', KATHRYN J. HOLM^, STEVEN J. PARR~ and NIRMAL J. SHAH' GEOLOGY, TOPOGRAPHY AND CLIMATE Bird is the northernmost island of the Seychelles, lying around 90 km north of Mahe, the largest of the granitic Seychelles, at the northern edge of the Seychelles bank. Different published sources vary in the estimated area of Bird Island with figures of c. 70 ha given by Feare (1979), 82 ha in Stoddart and Fosberg (1981), 101 ha in Skerrett et al. (2001), and 120.7 ha from recent aerial photographs (Ministry of Land Use and Habitat, Seychelles, unpublished data). In part, this variation may be explained by seasonal or longer-term variations in the vegetated area of the island; Bird Island is relatively dynamic, experiencing considerable coastal changes over time (Feare, 1979). The maximum elevation is less than 4 m above sea level. Unlike the majority of islands on the Seychelles Bank, Bird has no exposed granite and it is entirely formed of reef-derived sands. The accumulation of guano on sand deposits has led to the formation of phosphatic sandstone over 26% of the island's surface (Baker, 1963). Phosphatic sandstone is concentrated in a central band; the island's coastal zone is entirely sandy. Most of the original guano has now been removed for export. The soils of Bird Island are of two main series; over the central phosphatic sandstone area, Jemo series soils (missing their upper layer of guano) occur. -

Diversity of Insect Pollinators on Pigeon Pea, Cajanus Cajan L. In
Journal of Entomology and Zoology Studies 2018; 6(6): 47-50 E-ISSN: 2320-7078 P-ISSN: 2349-6800 Diversity of Insect pollinators on Pigeon pea, JEZS 2018; 6(6): 47-50 © 2018 JEZS Cajanus cajan L. in Odisha Received: 06-09-2018 Accepted: 09-10-2018 D Padhy D Padhy, CR Satapathy and RN Mohapatra AICRP on Honeybees and Pollinators, Department of Abstract Entomology, College of Agriculture, OUAT, A field experiment was conducted to study the “Diversity of insect pollinators on Pigeon pea (Cajanus Bhubaneswar, Odisha, India cajan L.) in Odisha” during Kharif, 2017-2018 at the Entomology block in Central Research Station of Orissa University of Agriculture and Technology, Bhubaneswar, Odisha. The Pigeon pea crop attracted CR Satapathy eleven species of pollinators represented by 3 families viz., Apidae (45.5%), Megachilidae (36.4%) and AICRP on Honeybees and Vespidae (18.2%) of order hymenoptera. Species wise diversity indicated that Megachile lanata Fab. was Pollinators, Department of the most dominant one (29.1%) followed by Megachile disjuncta Fab. (20.5%), Tetragonula iridipennis Entomology, College of Smith (14.2%), Apis cerana indica Fab. (10.95%), Megachile bicolor Fab. (10.7%), Xylocopa latipes Agriculture, OUAT, Drury (8.5%), Megachile hera Bingham (2.1%), Eumenes spp. (1.7%), Xylocopa aestuans Lin. (1.2%), Bhubaneswar, Odisha, India Vespa tropica Lin. (0.8%) and Apis dorsata Fab. (0.2%). All the pollinators were active during early and mid-flowering stage except X. latipes which was more active on the late flowering stage. During various RN Mohapatra flowering stages (69-159 DAS) of the crop the number of species varied from 5-11 with the Simpson’s AICRP on Honeybees and diversity index of 0.602-0.897. -

A Systematic Literature Review
Tropical Ecology 58(1): 211–215, 2017 ISSN 0564-3295 © International Society for Tropical Ecology www.tropecol.com Diversity of native bees on Parkinsonia aculeata L. in Jammu region of North-West Himalaya UMA SHANKAR, D. P. ABROL*, DEBJYOTI CHATTERJEE & S. E. H. RIZVI Division of Entomology, Sher-e- Kashmir University of Agricultural Sciences & Technology, Faculty of Agriculture, Chatha Jammu – 180009, J&K, India Abstract: A study was conducted in Jammu region of Jammu and Kashmir State to determine the species composition and relative abundance of pollinators on Parkinsonia aculeata L., (Family Fabaceae) is a perennial flowering plant, growing as an avenue tree on roadsides. Parkinsonia flowers attracted 27 species of insects belonging to orders Hymenoptera, Diptera and Lepidoptera. They included Megachile bicolor (Fabricius), Megachile hera (Bingham), Megachile lanata (Fabricius), Megachile disjuncta (Fab.), Megachile cephalotes (Smith), Megachile badia (Fab.), Megachile semivestita (Smith), Megachile vigilans (Smith), Megachile relata (Fab.), Megachile femorata, Andrena sp., Amegilla zonata (Linnaeus), Amegilla confusa (Smith), Apis dorsata (Fab.), Apis cerana (Fab.), Apis florea (Fab.), Ceratina smaragdula (Fab.)., Xylocopa latipes (Drury), Nomia iridescens (Smith), Nomia curvipes (Fab.), and seven species of unidentified insects. Megachile bees were most abundant and constituted more than 95% of the insects visiting Parkinsonia aculeata flowers. Species diversity measured by Shannon Wiener index showed a high value of H' = 2.03, reflecting a diverse pollinator community in the area. The foragers of all the species were found to be most active between 11.00 and 15.00 hrs and the population of flower visitors declined thereafter. Information on diversity of native pollinators from disturbed habitats and their specific dependence on P. -

Pollinator Biodiversity in Uganda and in Sub- Sahara Africa: Landscape and Habitat Management Strategies for Its Conservation
International Journal of Biodiversity and Conservation Vol. 3(11), pp. 551-609, 19 October, 2011 Available online at http://www.academicjournals.org/IJBC ISSN 2141-243X ©2011 Academic Journals Full Length Research Paper Pollinator biodiversity in Uganda and in Sub- Sahara Africa: Landscape and habitat management strategies for its conservation M. B. Théodore MUNYULI1, 2 1Department of Biology, National Center for Research in Natural Sciences, CRSN-Lwiro, D.S. Bukavu, South-Kivu Province, Democratic Republic of Congo. 2Department of Environmental and Natural Resource Economics, Faculty of Natural Resources and Environmental Sciences, Namasagali Campus, Busitema University., P .0. Box. 236, Tororo, eastern Uganda. E-mail: [email protected], [email protected], [email protected] Tel: +256-757356901, +256-772579267, +243997499842. Accepted 9 July, 2011 Previous pollinator faunistic surveys conducted in 26 different sites indicated that farmlands of central Uganda supported more than 650 bee species, 330 butterfly species and 57 fly species. Most crop species grown in Uganda are pollinator-dependents. There is also a high dependency of rural communities on pollination services for their livelihoods and incomes. The annual economic value attributable to pollinating services delivered to crop production sector was estimated to be worth of US$0.49 billion for a total economic value of crop production of US$1.16 billion in Uganda. Despite the great contribution of pollinators to crop yields, there is still lack of knowledge of their -

Study of Biodiversity of Wasps and Bees in Kota, Rajasthan, India
Journal of Entomology and Zoology Studies 2020; 8(6): 807-810 E-ISSN: 2320-7078 P-ISSN: 2349-6800 Study of biodiversity of wasps and bees in Kota, www.entomoljournal.com JEZS 2020; 8(6): 807-810 Rajasthan, India © 2020 JEZS Received: 11-09-2020 Accepted: 18-10-2020 Dr. Kusum Dang and Deoteema Doi Dr. Kusum Dang Associate professor, Department Abstract of zoology, Govt. P.G. College, Kota, Rajasthan, India Studies on diversity and distribution of insects are pre- requisites for biodiversity conservation. Wasps and bees both play important roles in ecosystem. The present study forms a part of evaluation of Deoteema Doi hymenopteran fauna (wasps and bees) in selected areas of Kota. And provides information about a good Student, Department of zoology, assemblage of species of wasps and bees in Kota, Rajasthan. The present study was performed in two Govt. P.G. College, Kota, chosen sites. Site 1 was Government College Kota Campus, Kota and Site 2 was Herbal Park, Kota. The Rajasthan, India study was conducted from January 2019 to May 2019. A total of 11 species of wasps and bees belonging to 6 families were observed and identified during the study from both the sites. 9 of them were identified up to species level and 2 up to genus level. Out of 11 species identified, 4 were of bees and rest 7 were of wasps. Species were also classified as abundant, occasional and rare based on field observations. 4 species of them were abundant, 4 species were occasional and 3 species were rare. Results of college campus show that, among bees, Family Apidae was most dominant with highest number of individuals and Apis dorsata Fabricius was most abundant; and among wasps, Family Vespidae was most abundant and Polistes hebraeus was most abundant species. -

A Plant Ecological Study and Management Plan for Mogale's Gate Biodiversity Centre, Gauteng
A PLANT ECOLOGICAL STUDY AND MANAGEMENT PLAN FOR MOGALE’S GATE BIODIVERSITY CENTRE, GAUTENG By Alistair Sean Tuckett submitted in accordance with the requirements for the degree of MASTER OF SCIENCE in the subject ENVIRONMENTAL MANAGEMENT at the UNIVERSITY OF SOUTH AFRICA SUPERVISOR: PROF. L.R. BROWN DECEMBER 2013 “Like winds and sunsets, wild things were taken for granted until progress began to do away with them. Now we face the question whether a still higher 'standard of living' is worth its cost in things natural, wild and free. For us of the minority, the opportunity to see geese is more important that television.” Aldo Leopold 2 Abstract The Mogale’s Gate Biodiversity Centre is a 3 060 ha reserve located within the Gauteng province. The area comprises grassland with woodland patches in valleys and lower-lying areas. To develop a scientifically based management plan a detailed vegetation study was undertaken to identify and describe the different ecosystems present. From a TWINSPAN classification twelve plant communities, which can be grouped into nine major communities, were identified. A classification and description of the plant communities, as well as, a management plan are presented. The area comprises 80% grassland and 20% woodland with 109 different plant families. The centre has a grazing capacity of 5.7 ha/LSU with a moderate to good veld condition. From the results of this study it is clear that the area makes a significant contribution towards carbon storage with a total of 0.520 tC/ha/yr stored in all the plant communities. KEYWORDS Mogale’s Gate Biodiversity Centre, Braun-Blanquet, TWINSPAN, JUICE, GRAZE, floristic composition, carbon storage 3 Declaration I, Alistair Sean Tuckett, declare that “A PLANT ECOLOGICAL STUDY AND MANAGEMENT PLAN FOR MOGALE’S GATE BIODIVERSITY CENTRE, GAUTENG” is my own work and that all sources that I have used or quoted have been indicated and acknowledged by means of complete references. -

Wasps and Bees in Southern Africa
SANBI Biodiversity Series 24 Wasps and bees in southern Africa by Sarah K. Gess and Friedrich W. Gess Department of Entomology, Albany Museum and Rhodes University, Grahamstown Pretoria 2014 SANBI Biodiversity Series The South African National Biodiversity Institute (SANBI) was established on 1 Sep- tember 2004 through the signing into force of the National Environmental Manage- ment: Biodiversity Act (NEMBA) No. 10 of 2004 by President Thabo Mbeki. The Act expands the mandate of the former National Botanical Institute to include respon- sibilities relating to the full diversity of South Africa’s fauna and flora, and builds on the internationally respected programmes in conservation, research, education and visitor services developed by the National Botanical Institute and its predecessors over the past century. The vision of SANBI: Biodiversity richness for all South Africans. SANBI’s mission is to champion the exploration, conservation, sustainable use, appreciation and enjoyment of South Africa’s exceptionally rich biodiversity for all people. SANBI Biodiversity Series publishes occasional reports on projects, technologies, workshops, symposia and other activities initiated by, or executed in partnership with SANBI. Technical editing: Alicia Grobler Design & layout: Sandra Turck Cover design: Sandra Turck How to cite this publication: GESS, S.K. & GESS, F.W. 2014. Wasps and bees in southern Africa. SANBI Biodi- versity Series 24. South African National Biodiversity Institute, Pretoria. ISBN: 978-1-919976-73-0 Manuscript submitted 2011 Copyright © 2014 by South African National Biodiversity Institute (SANBI) All rights reserved. No part of this book may be reproduced in any form without written per- mission of the copyright owners. The views and opinions expressed do not necessarily reflect those of SANBI. -

Biosearch 2004 Report
Biosearch Nyika: Malawi 2004 Edited by Marianne J Overton FOREWORD Peter Overton It is ten years since the Biosearch Nyika project was first mooted and agreement with the Director of National Parks and Wildlife obtained for our exploration of the remoter parts of the Nyika National Park. Over this period the teams have focused mainly on the northern part of the park where patrolling has been very limited and our gathering of intelligence has been most helpful to the Nyika management. In 2004 we undertook the most challenging expedition to date, launched from the extreme north of the park at Uledi, a four-hour drive from Thazima. The team‟s first challenge was to cross the unbridged North Rukuru River with all their supplies. They then had to climb up the western escarpment of the Mpanda ridge to a point on the Mpero River, where they set up a Base Camp, from which to launch out on their surveys. The greatest achievement was to climb both Mpanda and Kawozya and discover the remote Bleak House, now derelict but offering stunning views over Lake Malawi and far beyond. At this point they could certainly claim to be in remote country since this old site is much talked about but very rarely seen by visitors. We have yet to have clear information about who built it, when and why. Perhaps it was a holiday „retreat‟ for Livingstonia or a staging post for missionaries who conducted business on the west of the Nyika National Park and into Zambia. In many ways this expedition was the pinnacle of logistical achievement. -

Bee Conservation in Sub-Saharan Africa and Madagascar: Diversity, Status and Threats Connal D
Bee conservation in Sub-Saharan Africa and Madagascar: diversity, status and threats Connal D. Eardley, Mary Gikungu, Michael P. Schwarz To cite this version: Connal D. Eardley, Mary Gikungu, Michael P. Schwarz. Bee conservation in Sub-Saharan Africa and Madagascar: diversity, status and threats. Apidologie, Springer Verlag, 2009, 40 (3), 10.1051/apido/2009016. hal-00892023 HAL Id: hal-00892023 https://hal.archives-ouvertes.fr/hal-00892023 Submitted on 1 Jan 2009 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Apidologie 40 (2009) 355–366 Available online at: c INRA/DIB-AGIB/EDP Sciences, 2009 www.apidologie.org DOI: 10.1051/apido/2009016 Review article Bee conservation in Sub-Saharan Africa and Madagascar: diversity, status and threats* Connal D. Eardley1,MaryGikungu2, Michael P. Schwarz3 1 Agricultural Research Council, Private Bag X134, Queenswood, 0121, Pretoria, South Africa 2 Zoology Department, National Museums of Kenya, PO Box 40658-00100, Nairobi, Kenya 3 School of Biological Sciences, Flinders University, GPO Box 2100, Adelaide, South Australia 5001, Australia Received 14 October 2008 – Revised 2 February 2009 – Accepted 4 February 2009 Abstract – Sub-Saharan Africa and Madagascar contain a wealth of bee diversity, with particularly high levels of endemicity in Madagascar. -

Recapitulation of Genus Dinogamasus Mite (Acri/Mesostimata:Laelapidae ) on Carpenter Bee from India, with Description of a New Species
JOURNAL OF CRITICAL REVIEWS ISSN- 2394-5125 VOL 7, ISSUE 3, 2020 RECAPITULATION OF GENUS DINOGAMASUS MITE (ACRI/MESOSTIMATA:LAELAPIDAE ) ON CARPENTER BEE FROM INDIA, WITH DESCRIPTION OF A NEW SPECIES R. B. Andhale1, Aswini Pai2, Kalpana Pai3, R.S. Pandit4 Entomology Research Lab, Centre for advanced studies, Department of Zoology, Savitribai Phule Pune University, Pune India1. Department of Biology, St. Lawrance University, New York, USA2. [email protected], [email protected] Abstract: Phoretic mite species belong to genus Dinogamasus collected from large carpenter bees. Description and illustration of this new phoretic mite base on adult female are presented. The genus of Dinogamasus has recapitulated in India after ninety years. Key word: Xylocopa, Taxonomy, Phoretic mites, bee parasite Introduction: Mites are most successful free living organism adapted in parasitic, terrestrial, and aquatic habitat with highly diverse group among the arachnid (Oconnor 1982). A number of bees species have various kind of association with mites such as mutualistic, parasitoid, cleptoparasitic and commensal etc. Xylocopa bee is associated with Dinogamasus mites (Vitzthum 1930). Out of these associations, phoresy is a well known example. These are free living but most of time facultative associated with invertebrates for phoresy (temporarily use large organism for transportation or spend entire life cycle with host) as deutonymphs family of astigmatids like laelapidae, Chetodactylidae, and Canestriniidae (Oconnor 1982). Senertia genus has about sixty species of deutonymphs collected from carpenter bee (Baker & Delfinado-Baker 1983; Fain 1981; Ramaraju and Mohanasundaram 2001). Senertia Oudemans, 1905, is belonging family of chaetodactylidae, which is largest group of phoretic mites associates with carpenter bees (Eickwort 1994).