Fundamental Differences in Diversity and Biosynthetic Pathways
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sea Cucumber Abundance, Diversity and Fisheries in Samoa; an Assessment of Lagoon Occurring Sea Cucumbers
2006-05-17 Sea cucumber abundance, diversity and fisheries in Samoa; an assessment of lagoon occurring sea cucumbers Part I: A wide approached survey to assess status of commercial beche-de-mer species in Samoa. & Part II: The subsistence and artisanal sea cucumber fishery, with particular focus on Stichopus horrens, in Samoa. B.G.H. Eriksson Preface This study was performed in Samoa from 2005-09-20 to 2005-12-20 and finalises my university studies at Uppsala University towards an M.Sc in Biology. The work presented in this paper came about after a series of events and I owe greatly to all of those that are mentioned in the acknowledgement section. During 2005 a request was put forward to The Secretariat of the Pacific Community (SPC) from the Samoan Fisheries Division to perform a survey on the coastal resources (including sea cucumber resources) around the country of Samoa in the South Pacific. The coastal component of the Pacific Region Oceanic and Coastal Fisheries (PROCFish/C) section of SPC started up this work in collaboration with the Samoan Fisheries Division in June and August 2005 and covered finfish and invertebrate resources in parts of Upolu and Savaii. The invertebrate surveys included fisheries dependent and fisheries independent data collection. The fisheries independent surveys were in-water assessments of stock and habitat in grounds that was pre-selected because of fishing activities in that area. The data collected was generally density estimates (incl. species composition) across shifting habitats, but also biological data, such as length and weight measurements. Alongside this information fishery dependent data was also collected. -

Predator Defense Mechanisms in Shallow Water Sea Cucumbers (Holothuroidea)
PREDATOR DEFENSE MECHANISMS IN SHALLOW WATER SEA CUCUMBERS (HOLOTHUROIDEA) JESSICA A. CASTILLO Environmental Science Policy and Management, University of California, Berkeley, California 94720 USA Abstract. The various predator defense mechanisms possessed by shallow water sea cucumbers were surveyed in twelve different species and morphs. While many defense mechanisms such as the presence of Cuverian tubules, toxic secretions, and unpalatability have been identified in holothurians, I hypothesized that the possession of these traits as well as the degree to which they are utilized varies from species to species. The observed defense mechanisms were compared against a previously-derived phylogeny of the sea cucumbers of Moorea. Furthermore, I hypothesized that while the presence of such structures is most likely a result of the species’ placement on a phylogenetic tree, the degree to which they utilize such structures and their physical behavior are influenced by their individual ecologies. The presence of a red liquid secretion was restricted to individuals of the genus Holothuria (Linnaeus 1767) however not all members of the genus exhibited this trait. With the exception of H. leucospilota, which possessed both Cuverian tubules and a red secretion, Cuverian tubules were observed in members of the genus Bohadschia (Ostergren 1896). In accordance with the hypothesis, both the phylogenetics and individual ecology appear to influence predator defense mechanisms. However, even closely related species of similar ecology may differ considerably. Key words: holothurians; defense; toxicity; Cuverian tubules; Moorea, French Polynesia INTRODUCTION (Sakthivel et. Al, 1994). Approximately 20 species in two families and five genera, Sea cucumbers belong to the phylum including Holothuria, Bohadschia, and Thenelota, Echinodermata and the class Holothuroidea. -

High-Value Components and Bioactives from Sea Cucumbers for Functional Foods—A Review
Mar. Drugs 2011, 9, 1761-1805; doi:10.3390/md9101761 OPEN ACCESS Marine Drugs ISSN 1660-3397 www.mdpi.com/journal/marinedrugs Review High-Value Components and Bioactives from Sea Cucumbers for Functional Foods—A Review Sara Bordbar 1, Farooq Anwar 1,2 and Nazamid Saari 1,* 1 Faculty of Food Science and Technology, Universiti Putra Malaysia, Serdang, Selangor 43400, Malaysia; E-Mails: [email protected] (S.B.); [email protected] (F.A.) 2 Department of Chemistry and Biochemistry, University of Agriculture, Faisalabad 38040, Pakistan * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +60-389-468-385; Fax: +60-389-423-552. Received: 3 August 2011; in revised form: 30 August 2011 / Accepted: 8 September 2011 / Published: 10 October 2011 Abstract: Sea cucumbers, belonging to the class Holothuroidea, are marine invertebrates, habitually found in the benthic areas and deep seas across the world. They have high commercial value coupled with increasing global production and trade. Sea cucumbers, informally named as bêche-de-mer, or gamat, have long been used for food and folk medicine in the communities of Asia and Middle East. Nutritionally, sea cucumbers have an impressive profile of valuable nutrients such as Vitamin A, Vitamin B1 (thiamine), Vitamin B2 (riboflavin), Vitamin B3 (niacin), and minerals, especially calcium, magnesium, iron and zinc. A number of unique biological and pharmacological activities including anti-angiogenic, anticancer, anticoagulant, anti-hypertension, anti-inflammatory, antimicrobial, antioxidant, antithrombotic, antitumor and wound healing have been ascribed to various species of sea cucumbers. Therapeutic properties and medicinal benefits of sea cucumbers can be linked to the presence of a wide array of bioactives especially triterpene glycosides (saponins), chondroitin sulfates, glycosaminoglycan (GAGs), sulfated polysaccharides, sterols (glycosides and sulfates), phenolics, cerberosides, lectins, peptides, glycoprotein, glycosphingolipids and essential fatty acids. -

Pearlfish Carapus Bermudensis from the Sea Cucumber Holothuria Mexicana in Belize (Central America)
SPC Beche-de-mer Information Bulletin #38 – March 2018 73 Pearlfish Carapus bermudensis from the sea cucumber Holothuria mexicana in Belize (Central America) Arlenie Rogers1,*, Jean-François Hamel2 and Annie Mercier3 Pearlfish (Carapidae) are specialised fishes that mainly live in the respiratory tree of sea cucumber hosts (Arnold 1956; Shen and Yeh 1987; Smith and Tyler 1969; Smith 1964) in a relationship that has generally been defined as commensalism (Parmentier et al. 2003; Van Den Spiegel and Jangoux 1989; Parmentier et al. 2016). However, some species such as Encheliophis spp. are known to feed off their host’s gonad (Murdy and Cowan 1980; Parmentier et al. 2003; Pamentier and Vandewalle 2005; Parmentier et al. 2016). The present article highlights the occurrence of and from the Range (16˚05.616’N: 88˚42.827’W) on the pearlfish Carapus bermudensis (Figure 1) inside 12 February 2012 at a depth of 7.6 m. The latter the sea cucumber Holothuria mexicana in Belize. two sites consisted of seagrass (Thalassia testudi- Adults of H. mexicana were collected from Buggle num), sand and coral rubble and were within the Caye (16˚28.377’ N: 88˚21.77’W) on 14 July 2015 at Port Honduras Marine Reserve, while the former a depth 1.2 m; at Frenchman Caye (16˚06.347’N: site consisted of patch coral, sand and T. testudi- 88˚33.702’W) on 9 June 2014 at a depth of 10.7 m; num (Figure 1). Figure 1. Locations where sea cucumbers (H. mexicana) hosting the pearlfish C. bermudensis were found. -

1 Conference of the Parties to The
Conference of the Parties to the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES); Seventeenth Regular Meeting: Taxa Being Considered for Amendments to the CITES Appendices The United States, as a Party to the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), may propose amendments to the CITES Appendices for consideration at meetings of the Conference of the Parties. The seventeenth regular meeting of the Conference of the Parties to CITES (CoP17) is scheduled to be held in South Africa, September 24 to October 5, 2016. With this notice, we describe proposed amendments to the CITES Appendices (species proposals) that the United States might submit for consideration at CoP17 and invite your comments and information on these proposals. Please note that we published an abbreviated version of this notice in the Federal Register on August 26, 2015, in which we simply listed each species proposal that the United States is considering for CoP17, but we did not describe each proposal in detail or explain the rationale for the tentative U.S. position on each species. CITES is an international treaty designed to control and regulate international trade in certain animal and plant species that are affected by trade and are now, or potentially may become, threatened with extinction. These species are listed in the Appendices to CITES, which are available on the CITES Secretariat’s website at http://www.cites.org/sites/default/files/eng/app/2015/E-Appendices-2015-02-05.pdf. Currently, 181 Parties, including the United States, have joined CITES. -
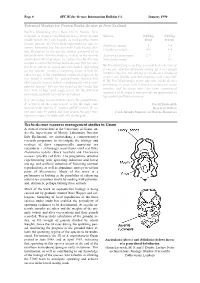
Beche-De-Mer Resource Management Studies in Guam
Page 8 SPC Bêche-de-mer Information Bulletin # 1 January 1990 Potential Market for Frozen Beche-de-mer in New Zealand By-Pro Marketing (P.O. Box 38679, Petone, New Zealand), is trying to establish itself as a beche-de-mer Species NZ$/kg NZ$/kg middleman in the Cook Islands, as well as other South dried frozen Pacific nations. By-Pro's field representative and co- Thelenota ananas 5 3-4 owner, Raymond Joe, has met with Cook Islands Ma- Holothuria nobilis 4-5 3-4 rine Resources to discuss the market potential of its beche-de-mer. Besides wanting to deal in the normal Actinopyga mauritiana 2-3 2 smoked/sun dried product, he claims that By-Pro has Bohadschia argus 2-3 located a market for frozen beche-de-mer that has sim- By-Pro Marketing is seeking as much beche-de-mer as ply been gutted, or gutted and then boiled, depending it can get, and has proposed setting up a sea freight on the species. Another interesting claim is that be- container/freezer, for storing beche-de-mer produced sides for any of the established commercial species, he in the Cook Islands, and then shipping it off when full. has found a market for gutted/frozen leopard fish If By-Pro Marketing's claim pan out, beche-de-mer (Bohadschia argus), which is traditionally not a com- processing in areas with freezers may become much mercial species. The species found in the Cooks that simpler, and for those who also have commercial they wish to buy, and rough prices for the different quantities of B. -

SPC Beche-De-Mer Information Bulletin
Secretariat of the Pacific Community ISSN 1025-4943 Issue 34 – May 2014 BECHE-DE-MER information bulletin Inside this issue Editorial The IUCN Red List assessment of th aspidochirotid sea cucumbers and its The 34 issue of the Beche-de-mer Information Bulletin includes, as implications always, a considerable amount of information on the biology, ecology and C. Conand et al. p. 3 bio-management of sea cucumbers. The status of the sea cucumber fishery in Batiki District, Lomaiviti, Fiji In the first article, Chantal Conand and co-authors describe the process used W. Lalavanua, I. Tuinasavusavu and the results obtained in an assessment of sea cucumber species for the and P. Seru p. 8 International Union for Conservation of Nature (IUCN) Red List; 16 threatened species, out of 377 known aspidochirotids examined, are presented. An Indonesian sea cucumber fishing village: The case of Pulau Misa The second article comes from Fiji. Watisoni Lalavanua and colleagues P.G. Navarro et al. p. 14 undertook a sea cucumber assessment survey in Batiki District in October An assessment of holothurian diversity, 2012. The results indicate that the sea cucumber fishery there is under abundance and distribution in the shallow lagoons of Mauritius stress from overexploitation and requires effective management. K. Lampe-Ramdoo, R. Moothien Pillay Pablo Navarro and co-authors provide some information on beche-de- and C. Conand p. 17 mer activities at Pulau Misa, a small island in Indonesia’s Flores Sea. The Some data on the diversity and people from Pulau Misa carry out a semi-traditional sea cucumber fishery. -

On the Association Between the Crab Hapalonotus Reticulatus (Crustacea
BULLETIN DE L'INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE, BIOLOGIE, 62: 167-177, 1992 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN, BIOLOGIE, 62: 167-177, 1992 On the association between the crab Hapalonotus reticulatus (Crustacea, Brachyura, Eumedonidae) and the sea cucumber Holothuria (Metriatyla) scabra (Echinodermata, Holothuridae) * by D. VANDENSPIEGEL, A. OVAERE & Cl. MASSIN Abstract Portunidae (genus Lissocarcinus) (for a review see J a n - g o u x , 1987). The Portunidae are exosymbionts, which A study of the fauna associated with holothurians from Hansa Bay live on the integument of the holothurian, while the (Papua New Guinea) reveals a new symbiotic association between the Pinnotheridae are endosymbionts, which generally live holothuroid Holothuria (Metriatyla) scabra and the crab Hapalonotus reticulatus. All the collected holothurians were infested, but with only in the gut or in the respiratory trees of their hosts. one crab per holothurian, always in the right respiratory tree near the Previously Hapalonotus reticulatus (D e M a n , 1879) was cloaca. Because of its size, H. reticulatus considerably expands the known from only 3 female specimens (with membrana wall of the respiratory tree, forming a membranaceous cyst. This ceous carapaces) from Ambon and Banda. Nothing is association, therefore, appears to be parasitic. However, the crab does not feed upon host tissue but seems to filter its food from water mentioned in the literature about the ecology of this passing through the respiratory tree of its host. crab. Only BALSS (1933) suggested that the body shape The description of two males allows to comment upon the taxonomical of the crab could be the result of a commensal way of position of H. -

Chemical Defense Mechanisms and Ecological Implications of Indo-Pacific Holothurians
molecules Article Chemical Defense Mechanisms and Ecological Implications of Indo-Pacific Holothurians Elham Kamyab 1,* , Sven Rohde 1 , Matthias Y. Kellermann 1 and Peter J. Schupp 1,2,* 1 Institute for Chemistry and Biology of the Marine Environment (ICBM), Carl-von-Ossietzky University Oldenburg, Schleusenstrasse 1, 26382 Wilhelmshaven, Germany; [email protected] (S.R.); [email protected] (M.Y.K.) 2 Helmholtz Institute for Functional Marine Biodiversity, University of Oldenburg, Ammerländer Heerstrasse 231, D-26129 Oldenburg, Germany * Correspondence: [email protected] (E.K.); [email protected] (P.J.S.); Tel.: +49-4421-944-100 (P.J.S.) Academic Editor: David Popovich Received: 14 August 2020; Accepted: 13 October 2020; Published: 19 October 2020 Abstract: Sea cucumbers are slow-moving organisms that use morphological, but also a diverse combination of chemical defenses to improve their overall fitness and chances of survival. Since chemical defense compounds are also of great pharmaceutical interest, we pinpoint the importance of biological screenings that are a relatively fast, informative and inexpensive way to identify the most bioactive organisms prior to further costly and elaborate pharmacological screenings. In this study, we investigated the presence and absence of chemical defenses of 14 different sea cucumber species from three families (Holothuriidae, Stichopodidae and Synaptidae) against ecological factors such as predation and pathogenic attacks. We used the different sea cucumber crude extracts as well as purified fractions and pure saponin compounds in a portfolio of ecological activity tests including fish feeding assays, cytotoxicity tests and antimicrobial assays against environmental pathogenic and non-pathogenic bacteria. -

Mechanisms Involved in Pearlfish Resistance to Holothuroid Toxins
Mar Biol (2016) 163:129 DOI 10.1007/s00227-016-2901-3 ORIGINAL PAPER Mechanisms involved in pearlfish resistance to holothuroid toxins Lola Brasseur1 · Eric Parmentier2 · Guillaume Caulier1 · Maryse Vanderplanck3 · Denis Michez3 · Patrick Flammang1 · Pascal Gerbaux4 · Georges Lognay5 · Igor Eeckhaut1 Received: 28 October 2015 / Accepted: 15 April 2016 / Published online: 4 May 2016 © Springer-Verlag Berlin Heidelberg 2016 Abstract Holothuroids produce triterpenoid saponins that against the toxin constituted by a larger secretion of mucus act as chemical defenses against predators and parasites. than other fishes, (2) a bioactive barrier against the tox- These saponins interact with sterols of the plasma mem- ins constituted by a mucus effective on saponins and (3) a branes, inducing the formation of pores and then cell lysis. Δ7sterol tissue composition mimicking holothuroids that To avoid such harms from their own saponins, holothuroids enable them to resist to saponins. First experiments showed present specific sterols in their tissues. Despite the noxious that the mucus has no effective impact on saponin chemical cytotoxic effect of their chemical defenses, holothuroids structures. Mass spectrometry analyses showed that carap- host various associates that display specific adaptations to ids, similarly to non-symbiotic fishes but contrary to their resist to saponin toxicity. Among them, symbiotic carapid hosts, present a Δ5sterol tissue composition. However, two fishes (i.e., pearlfishes) are resistant to ichthyotoxic sapo- different procedures have shown that carapids produce six nins as they display no stress response and a survival time to ten times more mucus than control fishes, suggesting that 45 times longer than free-living fishes without any specific a great quantity of mucus can protect carapids from their gill adaptation. -

Natural Spawning Observations of Pearsonothuria Graeffei And
38 SPC Beche-de-mer Information Bulletin #18 – May 2003 Ramofafia, C. and Byrne, M. 2001. Assessment of recruitment model”. Biological Bulletin the ‘tubule recruitment model’ in three tropi- 192:17–27. cal Aspidochirote holothurians. SPC Beche-de- Smiley, S. 1988. The dynamics of oogenesis and the Mer Information Bulletin 15:13–16. annual ovarian cycle of Stichopus californicus Sewell, M.A. 1992. Reproduction of the temperate (Echinodermata: Holothuroidea). Biological aspidochirote Stichopus mollis (Echinodermata: Bulletin 175:79–93. Holothuroidea) in New Zealand. Ophelia Tuwo, A. and Conand, C. 1992. Reproductive biol- 35(2):103–121. ogy of the holothurian Holothuria forskali Sewell, M.A., Tyler, P.A., Young, C.M. and Conand, (Echinodermata). Journal of Marine Biology C. 1997. Ovarian development in the class Association, United Kingdom 72:745–758. Holothuroidea: A reassessment of the “tubules Natural spawning observations of Pearsonothuria graeffei Pradina Purwati1 Spawning of Pearsonothuria graeffei in its natural habitat was observed during Anambas Expedition 2001. The animals stood up, waving the anterior part of the body slowly, and spilled their gametes into the water column. Dates and locations: 13 March 2002 on the north- eastern part of Jebung Bay, Jemaja Island (03°15.19'N and 106°13.48'E) and 14 March 2002 on the south western coast of Matak Island (02°52.43'N to 02°54.63'N and 105°50.97'E), Anambas islands, South China Sea. Depth: down to 32 m Time of spawning observation: 05:00–06:00 pm Habitat: reef slope, white sand, a lot of boulders and branching corals (first site); shallow dis- turbed fringing reef, damage and dead corals (second site). -

Molecular Phylogeny of Coral-Reef Sea Cucumbers (Holothuriidae: Aspidochirotida) Based on 16S Mitochondrial Ribosomal DNA Sequence
Molecular Phylogeny of Coral-Reef Sea Cucumbers (Holothuriidae: Aspidochirotida) Based on 16S Mitochondrial Ribosomal DNA Sequence Alexander M. Kerr,1* Daniel A. Janies,2, Ronald M. Clouse,2,à Yves Samyn,3 Jeni Kuszak,2 Junhyong Kim1,§ 1Department of Ecology and Evolutionary Biology, Osborn Zoo¨ logical Laboratories, Yale University, New Haven, CT 06520, U.S.A. 2Department of Invertebrate Zoology, American Museum of Natural History, New York, NY 10024, U.S.A. 3Global Taxonomy Initiative, Royal Belgian Institute of Natural Sciences, Vaurierstraat 29, B-1000 Brussels, Belgium Received: 4 February 2004 / Accepted: 5 May 2004 / Online publication: 30 December 2004 Abstract that, in addition to a probable bout of elaboration in ossicle form (the modification of rods or rosettes to Members of the Holothuriidae, found globally at low holothuriid-type buttons), at least 2 rounds of ossicle to middle latitudes, are often a dominant component simplification also transpired in which buttons re- of Indo–West Pacific coral reefs. We present the first verted to rods or rosettes. Cuvierian tubules, defen- phylogeny of the group, using 8 species from the 5 sive organs unique to numerous members of currently recognized genera and based on approxi- Holothuriidae, were probably present before the mately 540 nucleotides from a polymerase chain initial radiation of the family, but the reconstruction reaction–amplified and conserved 3¢ section of 16S is ambiguous as to their ancestral function. mitochondrial ribosomal DNA. Parsimony and likelihood analyses returned identical topologies, Key words: phylogeny — 16S rDNA — mtDNA — permitting several robust inferences to be drawn. Holothuriidae — Isostichopus — Stichopus Several points corroborated the Linnean classifica- tion.