Increasing Levels and Biomagnification of Persistent Organic Pollutants
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Politics of Roman Memory in the Age of Justinian DISSERTATION Presented in Partial Fulfillment of the Requirements for the D
The Politics of Roman Memory in the Age of Justinian DISSERTATION Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Marion Woodrow Kruse, III Graduate Program in Greek and Latin The Ohio State University 2015 Dissertation Committee: Anthony Kaldellis, Advisor; Benjamin Acosta-Hughes; Nathan Rosenstein Copyright by Marion Woodrow Kruse, III 2015 ABSTRACT This dissertation explores the use of Roman historical memory from the late fifth century through the middle of the sixth century AD. The collapse of Roman government in the western Roman empire in the late fifth century inspired a crisis of identity and political messaging in the eastern Roman empire of the same period. I argue that the Romans of the eastern empire, in particular those who lived in Constantinople and worked in or around the imperial administration, responded to the challenge posed by the loss of Rome by rewriting the history of the Roman empire. The new historical narratives that arose during this period were initially concerned with Roman identity and fixated on urban space (in particular the cities of Rome and Constantinople) and Roman mythistory. By the sixth century, however, the debate over Roman history had begun to infuse all levels of Roman political discourse and became a major component of the emperor Justinian’s imperial messaging and propaganda, especially in his Novels. The imperial history proposed by the Novels was aggressivley challenged by other writers of the period, creating a clear historical and political conflict over the role and import of Roman history as a model or justification for Roman politics in the sixth century. -

ROTEX Gassolarunit Gas Condensing Boiler with Stratified Solar Storage Tank
For specialist technical operation ROTEX GasSolarUnit Gas condensing boiler with stratified solar storage tank Installation and maintenance instructions 0085 BM 0065 Type Rated thermal output GB ROTEX GSU 320 3 - 20 kW modulating Edition 09/2007 ROTEX GSU 520S 3 - 20 kW modulating ROTEX GSU 530S 7 - 30 kW modulating ROTEX GSU 535 8 - 35 kW modulating Manufacture number Customer Guarantee and conformity ROTEX accepts the guarantee for material and manufacturing defects according to this statement. Within the guarantee period, ROTEX agrees to have the device repaired by a person assigned by the company, free of charge. ROTEX reserves the right to replace the device. The guarantee is only valid if the device has been used properly and it can be proved that it was installed properly by an expert firm. As proof, we strongly recommend completing the enclosed installation and instruction forms and returning them to ROTEX. Guarantee period The guarantee period begins on the day of installation (billing date of the installation company), however at the latest 6 months after the date of manufacture (billing date). The guarantee period is not extended if the device is returned for repairs or if the device is replaced. Guarantee period of burner, boiler body and boiler electronics: 2 years Guarantee exclusion Improper use, intervention in the device and unprofessional modifications immediately invalidate the guarantee claim. Dispatch and transport damage are excluded from the guarantee offer. The guarantee explicitly excludes follow-up costs, especially the assembly and disassembly costs of the device. There is no guarantee claim for wear parts (according to the manufacturer's definition), such as lights, switches, fuses. -

Jordanes and the Invention of Roman-Gothic History Dissertation
Empire of Hope and Tragedy: Jordanes and the Invention of Roman-Gothic History Dissertation Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Brian Swain Graduate Program in History The Ohio State University 2014 Dissertation Committee: Timothy Gregory, Co-advisor Anthony Kaldellis Kristina Sessa, Co-advisor Copyright by Brian Swain 2014 Abstract This dissertation explores the intersection of political and ethnic conflict during the emperor Justinian’s wars of reconquest through the figure and texts of Jordanes, the earliest barbarian voice to survive antiquity. Jordanes was ethnically Gothic - and yet he also claimed a Roman identity. Writing from Constantinople in 551, he penned two Latin histories on the Gothic and Roman pasts respectively. Crucially, Jordanes wrote while Goths and Romans clashed in the imperial war to reclaim the Italian homeland that had been under Gothic rule since 493. That a Roman Goth wrote about Goths while Rome was at war with Goths is significant and has no analogue in the ancient record. I argue that it was precisely this conflict which prompted Jordanes’ historical inquiry. Jordanes, though, has long been considered a mere copyist, and seldom treated as an historian with ideas of his own. And the few scholars who have treated Jordanes as an original author have dampened the significance of his Gothicness by arguing that barbarian ethnicities were evanescent and subsumed by the gravity of a Roman political identity. They hold that Jordanes was simply a Roman who can tell us only about Roman things, and supported the Roman emperor in his war against the Goths. -

The Aksumites in South Arabia: an African Diaspora of Late Antiquity
Chapter 11 The Aksumites in South Arabia: An African Diaspora of Late Antiquity George Hatke 1 Introduction Much has been written over the years about foreign, specifically western, colo- nialism in sub-Saharan Africa, as well as about the foreign peoples, western and non-western alike, who have settled in sub-Saharan Africa during the modern period. However, although many large-scale states rose and fell in sub- Saharan Africa throughout pre-colonial times, the history of African imperial expansion into non-African lands is to a large degree the history of Egyptian invasions of Syria-Palestine during Pharaonic and Ptolemaic times, Carthagin- ian (effectively Phoenician) expansion into Sicily and Spain in the second half of the first millennium b.c.e, and the Almoravid and Almohad invasions of the Iberian Peninsula during the Middle Ages. However, none of this history involved sub-Saharan Africans to any appreciable degree. Yet during Late Antiquity,1 Aksum, a sub-Saharan African kingdom based in the northern Ethi- opian highlands, invaded its neighbors across the Red Sea on several occasions. Aksum, named after its capital city, was during this time an active participant in the long-distance sea trade linking the Mediterranean with India via the Red Sea. It was a literate kingdom with a tradition of monumental art and ar- chitecture and already a long history of contact with South Arabia. The history of Aksumite expansion into, and settlement in, South Arabia can be divided into two main periods. The first lasts from the late 2nd to the late 3rd century 1 Although there is disagreement among scholars as to the chronological limits of “Late Antiq- uity”—itself a modern concept—the term is, for the purposes of the present study, used to refer to the period from ca. -

3.2 Precipitation Or Dry-Wet Reconstructions
Climate change in China during the past 2000 years: An overview Ge Quansheng , Zheng Jingyun Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China Email: [email protected] Outline 1 Introduction 2 Historical Documents as Proxy 3 Reconstructions and Analyses 4 Summary and Prospects 1. Introduction: E Asia2K Climate System Socio-economic System •typical East Asian •dense population and rapid monsoon climate economic development • significant seasonal and • be susceptible to global inter-annual and inter- warming and extreme decadal variability climate events Climate change study in the past 2ka in East Asian is both beneficial and advantageous. • various types of natural proxy • Plenty of historical documents Fig. Active regional working groups under as proxy the past 2ka theme (PAGES 2009) 2. Historical Documents as Proxy Type Period Amount Chinese classical 1,531 kinds, 137 BC~1470 AD documents 32,251 volumes More than 8,000 1471~1911 (The Ming Local gazettes books (部), 110, and Qing Dynasty) 000 volumes Memos to the About 120,000 1736~1911 emperor pieces Archives of the 1912~1949 20,000 volumes Republic of China More than 200 Private diaries 1550~ books (部) Chinese classical documents AD 833, North China plain: Extreme drought event was occurred, crops were shriveling, no yields, people were in hungry…. Fig. Example for Ancient Chinese writings Local gazettes The 28th year of the Daoguang reign (1848 AD), the 6th (lunar) month, strong wind and heavy rain, the Yangtze River overflowed; the 7th month, strong wind Fig. Gazettes of Yangzhou Prefecture and thunder storm, field published in 1874 AD and houses submerged. -

Objectives, Policy, and Scope of Operations Chapter 10
Objectives, Policy, and Scope of Operations Chapter 10 CHAPTER 10 OBJECTIVES, POLICY AND SCOPE OF OPERATION MISSION STATEMENT The Northwest Interagency Coordination Center (NWCC) serves all Federal, and cooperating State Wildland Fire Agencies in the Northwest Geographic Area. NWCC coordinates Interagency presuppression and suppression strategies and facilitates intelligence and logistical support related to existing and anticipated responses to all-risk incidents. The central focus of NWCC operation is Wildland Fire. As an agent of the NW Multi-Agency Coordinating Group (NWMAC), NWCC continually assesses the Geographic Area situation and, utilizing strategic intent, allocates or reallocates resources in coordination with agency duty officers, incident management teams, dispatch center managers, and local unit leadership for the purpose of sustaining effective emergency and/or prescribed fire operations. NWCC is the Geographical Area hub for out-of-area resource mobilizations, working in coordination with other Geographical Area Coordination Centers and the National Interagency Coordination Center located in Boise, Idaho. PRIORITIES To effectively manage resource competition, the NWCC Manager, Emergency Operations Manager, and/or the NWMAC will establish priorities for allocating resources to incidents within the Geographic Area. Refer to NMG 10, for specific criteria. LOCAL AND GEOGRAPHIC AREA DRAWDOWN LEVELS AND NATIONAL READY RESERVE Local drawdown is established by the local unit and/or the local MAC Group and implemented by the local dispatch office. The local dispatch office will notify NWCC of local drawdown decisions and actions. Geographic area drawdown is established by NWMAC and implemented by NWCC. NWCC will notify the local dispatch offices and the NICC of the Northwest drawdown decisions and actions. -
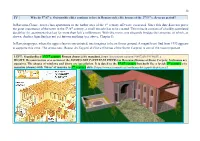
IV in Ravenna-Classe, Lower-Class Apartments in the Harbor Area of The
38 IV Why do 5th/6th c. Ostrogothic elites continue to live in Roman-style elite houses of the 2nd/3rd c. Severan period? In Ravenna-Classe, lower-class apartments in the harbor area of the 1st century AD were excavated. Since this date does not prove the great importance of the town in the 5th/6th century, a small miracle has to be created. This miracle consists of a boldly postulated durability for apartments that last for more than half a millennium. With this move, one elegantly bridges the centuries, of which, as shown, Andrea Agnellus has not yet known anything (see above, Chapter I). In Ravenna proper, where the upper class is concentrated, one imagines to be on firmer ground. A magnificent find from 1993 appears to supports this view. The aristocratic Domus dei Tappeti di Pietra (Domus of the Stone Carpets) is one of the most important LEFT: Standardized 1st/2nd century Roman domus (city mansion). [https://pl.pinterest.com/pin/91057223699970657/.] RIGHT: Reconstruction of a section of the DOMUS DEI TAPPETI DI PIETRA in Ravenna (Domus of Stone Carpets; bedrooms are upstairs). The shapes of windows and doors are speculation. It is dated to the 5th/6th century but built like a lavish 2nd century city mansion (domus) with 700 m2 of mosaics in 2nd century style. [https://www.ravennantica.it/en/domus-dei-tappeti-di-pietra-ra/.] 39 Italian archaeological sites discovered in recent decades. Located inside the eighteenth-century Church of Santa Eufemia, in a vast underground environment located about 3 meters below street level, it consists of 14 rooms paved with polychrome mosaics and marble belonging to a private building of the fifth-sixth century. -

I 2 1 8 1 9 20 2L 22 24 25 26 2L 28 29 BE IT FURTHER RESOLVED
------t I 2 RESOLUTION NO. O8-7 J -A A RESOLUTION OF THE ALBANY CITY COUNCIL ESTABLISHING THE COMMUNITY MEDIA ACCESS COMMITTEE 5 6 WHEREAS, the Albany City Council approvedthe conceptof creatinga Community l Media AccessCommittee fiom the ad hoc CableTelevision Committee on Februarv4. 8 2008;and 9 10 WHEREAS, the Community Media AccessCommittee will focuson Public Education 11 Government(PEG) cabletelevision access in Albanv: and I2 1a l-l NOW THEREFORE BE IT RESOLVED, that the Albany City Council hereby I4 establishesa CommunityMedia AccessCommittee to advisethe City Councilon cable l5 televisionrelated issues. l6 t7 BE IT FURTHER RESOLVED: 18 19 1. The Community Media AccessCommittee shall be composedof five 20 members,one eachappointed by Council members,with termsof two years, 2l effectiveJanuary, 2009. Until January2009, the membersof the ad-hocCable 22 TelevisionCommittee shall serveas the membersof the Community Media L) AccessCommittee. 24 25 Z. The chargeto the Committeeshall be to: 26 A. Preparefive-year operation plans for KALB 2l B. Advise City on changesto the CableTV accesspolicy 28 C. Evaluatecurrent goals, create new goals,and suggesttimelines for all goals 29 D. Overseefunds that are spenton cable-relatedactivities 'll E. Encouragealternative funding sourcesfor equipmentand staff residents 2 ll F. Provide video productiontraining opportunities for Albany il G. CreateAlbany-based programming for the channel and 3 ll post KALB programmingschedules in cabletelevision guide channels tl H. 4ll in other media I in cable-relatedactivities -l L Coordinateand recruit volunteersto participate 1% PEG J. Recommendthe allocationof fundsfrom the new statefranchise tlol suPPortfee KALB 7l K. Promoteand Publicize 8lI or I creatingthe committee, it is not the intent of the council to duplicate ol'l 3. -
Ancient Arabia a Brief History and Time-Line.Pages
Ancient Arabia: A brief history and time-line ! 1. Multiple Arabias Ancient sources, in particular the Assyrian Annals and the Greek and Roman historians and geographers, defined ‘Arabia’ as anywhere inhabited by peoples called ‘Arabs’. They applied this term to populations with very different ways-of-life in a wide variety of areas, and it seems likely that only a combination of a recognizably common language and certain common cultural traits could have defined them all as belonging to the same group. "1 From the 8th century BC, the Assyrians and Babylonians recorded Arabs living (from east to west) in eastern Mesopotamia between the Tigris and Iran, settled in large numbers in Babylonia, in the Syrian Jazīra (between the Tigris and Euphrates), on the slopes of the Anti-Lebanon mountains (between modern Lebanon and Syria), in north and north-west Arabia, and in Sinai. By the 6th century, the Achaemenid Persian empire recognized an Arab enclave in Gaza and its hinterland, and a century later Herodotus regarded ‘Arabia’ as being most of eastern Egypt, between the Nile and the Red Sea. Xenophon (c. 430–c. 354 BC) found ‘Arabias’ in northern Syria and northern and central Mesopotamia, while Alexander the Great (356–323 BC) encountered Arabs in the Lebanon and Anti-Lebanon mountains, Gaza, Sinai, the eastern Nile Delta, and eastern Egypt, as well as at the head of the Persian Gulf, and even in central Iran. His explorers also identified and defined the ‘Arabian’ Peninsula for the first time, and, by the Roman period, there were ‘Arabias’ in eastern Egypt and the delta, Sinai, southern Palestine (the Negev), the Beqaʿ Valley of Lebanon and the Anti- Lebanon, the whole of modern Jordan, southern, central and northern Syria, northern, central, and southern Mesopotamia, as well as the Peninsula. -
393 41. Numismatic Notes. SOME THOUGHTS on the HISTORY OF
41. Numismatic notes. SOME THOUGHTS ON THE HISTORY OF A CURIOUS REVERSE DIE1 Figure 1. Numismatic Circular, September 1981, no. 6111. Figure 2. Catalogue 35.13. Figure 3. Private collection. Students of the Byzantine series and many other collectors will be familiar with the follis type which has, as its obverse, a profile bust and, on the reverse, the denominational mark M with the mint signature CON below and a six or eight pointed star to either side. This design seems to have been struck continuously between circa 507 and 532 thereby straddling the reigns of Anastasius I, Justin I and Justinian I. A chance comparison between two coins, one each in the collection of two London Numismatic Club members, has provoked an intriguing problem. The coins are die-linked, both having been struck with a flawed reverse die which has imparted a pronounced mark above the right hand star on each example. What is therefore most interesting is that the obverse inscriptions bear the names of Anastasius I (figure 1) and Justinian I (figure 2) respectively. It would be exceptional, even in the context of the relaxed regime that frequently characterises Byzantine mint administration, if a reverse die had survived for a period of over 10 years and took in three reigns. The mint mark, however, actually reads NOC indicating that the maker correctly inverted the individual letters, but forgot to reverse their order when cutting the die. This, together with certain other features - retrograde letters belonging to the obverse inscriptions of both specimens 1 Donald, P. J., and Mansfield, S. -

Justinian and the Unity of Faith and Empire: the Continual Search for Compromise
JUSTINIAN AND THE UNITY OF FAITH AND EMPIRE The Continual Search for Compromise A thesis presented by ANDREW STEPHENSON to The School of Art History, Cinema, Classics and Archaeology in partial fulfilment of the requirements for the degree of Bachelor of Arts (Honours) in the subject area of Classical Studies The University of Melbourne SUPERVISOR: ASSOCIATE PROFESSOR ROGER SCOTT MAY, 2004 SYNOPSIS The Byzantine Emperor Justinian has been praised for his major building programme, his codification of the law and his reconquest of Africa and Italy, while at the same time being criticized for the protracted campaigns involved in the reconquest and for his continual meddling in Church affairs. But since the time of Constantine the unity of Church and State had been a major developing theme in the Roman Empire, particularly in the East. As well as being heir to this development, Justinian inherited a schism between the East and the Papacy in the West, as well as doctrinal division within the East itself. For over forty-five years Justinian strove to find a doctrinal solution that would satisfy the various factions and would bind the Empire together for its Christian security and salvation. His major achievement was to work continually towards this by focussing on the elucidation and clarification of doctrine through the unremitting application of his consummate skill. His failure to achieve lasting unity was not a fault of his but was due to the intransigence of the competing factions that he sought to unify. CONTENTS Introduction 1 1. Justinian: The Familiar Picture 3 2. The Emperor’s Heresy 7 3. -

Byzantium Becomes the New Rome
269-273-0311s1 10/11/02 3:53 PM Page 269 TERMS & NAMES 1 • Justinian Code Byzantium Becomes • Hagia Sophia • patriarch • icon • iconoclast the New Rome •excommunication • schism MAIN IDEA WHY IT MATTERS NOW • Cyrillic alphabet Constantinople ruled an eastern empire Byzantine culture deeply influenced that survived for over a thousand years. Orthodox Christianity, a major branch of modern Christianity. SETTING THE STAGE The western Roman Empire crumbled in the 5th century as it was overrun by invading Germanic tribes. (See Chapter 6.) The threat to the empire, however, was already apparent in the 4th century. Emperor Constantine rebuilt the old port city of Byzantium on the Bosporus strait for two reasons. In Byzantium, he could respond to the danger of the Germanic tribes. He could also be close to his rich eastern provinces. He renamed the city Constantinople and in the year 330, he made Both sides of this it the capital of the empire. gold medallion display Emperor Justinian as a A New Rome in a New Setting military commander. But the coin really Constantine planned Constantinople as the new capital of the empire—the New celebrates a victory Rome. As a result of his decision, the empire’s center of power moved eastward. The by General eastern provinces then began to develop independently of the declining West. An Belisarius. The emperor often eastern empire would gradually come into being. feared that the general’s popularity Justinian: A New Line of Caesars Because of the difficulties of communication would outshine his between the eastern and troubled western parts of the empire, they were officially own.