Innate and Learned Aspects of Pheromone-Mediated Social Behaviours
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ant Trails: a Key to Management with Baits1
ENY-259 Ant Trails: A Key to Management with Baits1 John Klotz, Dave Williams, Byron Reid, Karen Vail, and Phil Koehler2 Communication in the ants is based on chemical straight back to the nest (Figure 1). Somehow on the signals. These chemicals are called pheromones and outgoing trip she can keep track of her position with vary from alarm and nestmate recognition, to the one respect to her nest, and, on the return trip, uses this we will focus on here, recruitment. All of the pest information to take the shorter, more direct route ants use odor trails for orientation, but these trails home. On the way back to the nest, she lays down an differ from one species to another. Where the odor trail. Once back in the nest, this scout ant then pheromones originate in the ant's body, their alerts her nestmates of the food find, which chemical composition, as well as how long they last, encourages them to leave the nest. These recruited all vary from one ant species to the next. In fire ants, ants will follow the odor trail directly to the food the trail chemical is produced by the Dufour's gland, source. In turn, each ant will reinforce the odor trail which is named after its discoverer, Dufour, and is until the food is gone. This behavior is a highly laid down by the stinger. This pheromone is made up efficient means of exploiting a temporary food of molecules which evaporate very quickly. Thus, the resource. fire ant's odor trail is very short-lived. -

Medical Review Officer Manual
Department of Health and Human Services Substance Abuse and Mental Health Services Administration Center for Substance Abuse Prevention Medical Review Officer Manual for Federal Agency Workplace Drug Testing Programs EFFECTIVE OCTOBER 1, 2010 Note: This manual applies to Federal agency drug testing programs that come under Executive Order 12564 dated September 15, 1986, section 503 of Public Law 100-71, 5 U.S.C. section 7301 note dated July 11, 1987, and the Department of Health and Human Services Mandatory Guidelines for Federal Workplace Drug Testing Programs (73 FR 71858) dated November 25, 2008 (effective October 1, 2010). This manual does not apply to specimens submitted for testing under U.S. Department of Transportation (DOT) Procedures for Transportation Workplace Drug and Alcohol Testing Programs (49 CFR Part 40). The current version of this manual and other information including MRO Case Studies are available on the Drug Testing page under Medical Review Officer (MRO) Resources on the SAMHSA website: http://www.workplace.samhsa.gov Previous Versions of this Manual are Obsolete 3 Table of Contents Chapter 1. The Medical Review Officer (MRO)........................................................................... 6 Chapter 2. The Federal Drug Testing Custody and Control Form ................................................ 7 Chapter 3. Urine Drug Testing ...................................................................................................... 9 A. Federal Workplace Drug Testing Overview.................................................................. -

Mouse Breeding Colony Management 1. Mouse Reproduction A. General Mouse Information I. the Average Mouse Lives Approximately
Mouse Breeding Colony Management 1. Mouse Reproduction A. General Mouse Information i. The average mouse lives approximately 2.5 years; however, the reproductive life span of mice is significantly shorter at 7-8 months. ii. Most mice reach sexual maturity (males and females) at 4-7 weeks of age. Younger mice generally produce smaller litters and therefore are not typically mated until they reach 6-8 weeks, of age. Mice that have been housed alone or in same-sex pairs will usually not breed successfully if they are older than 6-8 months. iii. The mouse estrous cycle is 4-5 days in length. Mice cycle continuously throughout the year (non-seasonal breeders). Female mice are only receptive to males when they are in estrus. Mating typically occurs at night (lights off). Ovulation occurs 8-12 hours after the onset of estrous. iv. If fertilization occurs, fetuses can be palpated by day 14. v. Gestation in mice is typically 19-21 days (strain dependent). vi. Parturition in mice may last 1-3 hours and frequently occurs at night. Females will go into estrus within 24 hours of parturition and are sexually receptive during this time. vii. Litter size varies among strains, but averages 4-12 pups. Inbred mice tend to have smaller litters than outbred mice. viii. Mice are typically weaned at 21-28 days or at 10g of body weight. The Purdue Animal Care and Use Committee requires that mouse pups be weaned at 21 days unless PACUC approval is given on an approved animal use protocol. See Policy attached. -

Algal Sex Determination and the Evolution of Anisogamy James Umen, Susana Coelho
Algal Sex Determination and the Evolution of Anisogamy James Umen, Susana Coelho To cite this version: James Umen, Susana Coelho. Algal Sex Determination and the Evolution of Anisogamy. Annual Review of Microbiology, Annual Reviews, 2019, 73 (1), 10.1146/annurev-micro-020518-120011. hal- 02187088 HAL Id: hal-02187088 https://hal.sorbonne-universite.fr/hal-02187088 Submitted on 17 Jul 2019 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Annu. Rev. Microbiol. 2019. 73:X–X https://doi.org/10.1146/annurev-micro-020518-120011 Copyright © 2019 by Annual Reviews. All rights reserved Umen • Coelho www.annualreviews.org • Algal Sexes and Mating Systems Algal Sex Determination and the Evolution of Anisogamy James Umen1 and Susana Coelho2 1Donald Danforth Plant Science Center, St. Louis, Missouri 63132, USA; email: [email protected] 2Sorbonne Université, UPMC Université Paris 06, CNRS, Algal Genetics Group, UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, CS 90074, F-29688, Roscoff, France [**AU: Please write the entire affiliation in French or write it all in English, rather than a combination of English and French**] ; email: [email protected] Abstract Algae are photosynthetic eukaryotes whose taxonomic breadth covers a range of life histories, degrees of cellular and developmental complexity, and diverse patterns of sexual reproduction. -

Behavioral Roles of Oxytocin and Vasopressin
Chapter 3 Behavioral Roles of Oxytocin and Vasopressin Benjamin C. Nephew Additional information is available at the end of the chapter http://dx.doi.org/10.5772/50422 1. Introduction Arginine Vasopressin (AVP) and oxytocin (OXT) are peptide hormones found in most mammals that have vital physiological and behavioral actions. The major sites of AVP production are the paraventricular (PVN) and supraoptic (SON) nuclei in the hypothalamus, although AVP and its receptors are found in numerous brain nuclei and peripheral tissues. AVP’s physiological roles, which are mediated through both peripheral and central mechanisms, include regulating fluid homeostasis and blood pressure. It is also an important component of the endocrine stress response through its actions in the posterior pituitary gland, where it is a secretagogue of ACTH, stimulating the release of corticosteroid stress hormones and catecholamines from the adrenal glands. The three receptor subtypes for AVP are V1a, V1b, and V2. V2 receptors mediate the fluid regulating actions of AVP in the periphery, where the behavioral and central endocrine functions of AVP are mediated by the V1a and V1b receptors in the brain. These receptors are also involved in the central control of cardiovascular activity. Oxytocin’s major physiological roles are to facilitate uterine contractions during birth through a positive feedback mechanism during the second and third stages of labor, and to mediate milk letdown. In lactating mammalian mothers, OXT initiates milk letdown in the mammary glands, and the release of OXT is stimulated by suckling. OXT has one known receptor which has several alleles. The focus of the present chapter will be on the social behavior functions of both AVP and OXT. -

Color, Taste, and Odor: What You Should Know
Color, Taste, and Odor: What you should know From time to time the MassDEP receives consumer questions or complaints regarding the look, taste or the odor of drinking water. Listed below are common problems with drinking water and their most common causes. Please note that a particular problem in your drinking water may be the result of a cause not listed here; the only way to confirm a cause is to have a certified lab analyze the water and discuss the results with drinking water professional. If you receive water from a public drinking water system it is important to contact the Public Water Supply (PWS) before having a laboratory analyze the water. Information on private water testing is available. Filtering or treating the water may remedy persistent problems; however MassDEP does not recommend filtering or treating your water supply if your water is supplied by a MassDEP- approved PWS. MassDEP also does not regulate or recommend specific treatment systems for private home use. If you decide to use a filtration or treatment device in your home, the Department strongly encourages you to contact National Sanitation Foundation (NSF) for a list of approved devices. If you purchase a treatment device for private home use MassDEP also strongly recommends that it is maintained and provide active maintenance according to the manufacturer's instructions. Failure to maintain the equipment properly may make treatment ineffective and/or may create the potential for contamination. Common problems with drinking water are grouped into three categories: Color problems Taste / odor problems Particles in water If the problem with your water is not described here, if you are on a public water system please contact the public water department in your city or town or the MassDEP Drinking Water Program at your nearest regional MassDEP office. -

Mice in a Labyrinth Show Rapid Learning, Sudden Insight, and Efficient Exploration Matthew Rosenberg1†, Tony Zhang1†, Pietro Perona2*, Markus Meister1*
RESEARCH ARTICLE Mice in a labyrinth show rapid learning, sudden insight, and efficient exploration Matthew Rosenberg1†, Tony Zhang1†, Pietro Perona2*, Markus Meister1* 1Division of Biology and Biological Engineering, California Institute of Technology, Pasadena, United States; 2Division of Engineering and Applied Science, California Institute of Technology, Pasadena, United States Abstract Animals learn certain complex tasks remarkably fast, sometimes after a single experience. What behavioral algorithms support this efficiency? Many contemporary studies based on two-alternative-forced-choice (2AFC) tasks observe only slow or incomplete learning. As an alternative, we study the unconstrained behavior of mice in a complex labyrinth and measure the dynamics of learning and the behaviors that enable it. A mouse in the labyrinth makes ~2000 navigation decisions per hour. The animal explores the maze, quickly discovers the location of a reward, and executes correct 10-bit choices after only 10 reward experiences — a learning rate 1000-fold higher than in 2AFC experiments. Many mice improve discontinuously from one minute to the next, suggesting moments of sudden insight about the structure of the labyrinth. The underlying search algorithm does not require a global memory of places visited and is largely explained by purely local turning rules. Introduction How can animals or machines acquire the ability for complex behaviors from one or a few experien- ces? Canonical examples include language learning in children, where new words are learned after *For correspondence: just a few instances of their use, or learning to balance a bicycle, where humans progress from com- [email protected] (PP); plete incompetence to near perfection after crashing once or a few times. -

Pheromone-Based Disruption of Eucosma Sonomana and Rhyacionia Zozana (Lepidoptera: Tortricidae) Using Aerially Applied Microencapsulated Pheromone1
361 Pheromone-based disruption of Eucosma sonomana and Rhyacionia zozana (Lepidoptera: Tortricidae) using aerially applied microencapsulated pheromone1 Nancy E. Gillette, John D. Stein, Donald R. Owen, Jeffrey N. Webster, and Sylvia R. Mori Abstract: Two aerial applications of microencapsulated pheromone were conducted on five 20.2 ha plots to disrupt western pine shoot borer (Eucosma sonomana Kearfott) and ponderosa pine tip moth (Rhyacionia zozana (Kearfott); Lepidoptera: Tortricidae) orientation to pheromones and oviposition in ponderosa pine plantations in 2002 and 2004. The first application was made at 29.6 g active ingredient (AI)/ha, and the second at 59.3 g AI/ha. Baited sentinel traps were used to assess disruption of orientation by both moth species toward pheromones, and E. sonomana infes- tation levels were tallied from 2001 to 2004. Treatments disrupted orientation by both species for several weeks, with the first lasting 35 days and the second for 75 days. Both applications reduced infestation by E. sonomana,but the lower application rate provided greater absolute reduction, perhaps because prior infestation levels were higher in 2002 than in 2004. Infestations in treated plots were reduced by two-thirds in both years, suggesting that while increas- ing the application rate may prolong disruption, it may not provide greater proportional efficacy in terms of tree pro- tection. The incidence of infestations even in plots with complete disruption suggests that treatments missed some early emerging females or that mated females immigrated into treated plots; thus operational testing should be timed earlier in the season and should comprise much larger plots. In both years, moths emerged earlier than reported pre- viously, indicating that disruption programs should account for warmer climates in timing of applications. -

The Study of Animal Behaviour and Its Applications
Eth. Dom. Animals - Chap 01 22/4/02 9:48 am Page 3 The Study of Animal Behaviour 1 and its Applications Per Jensen Introduction Ethology is the science whereby we study animal behaviour, its causa- tion and its biological function. But what is behaviour? If we spend a few minutes thinking about this, a number of answers may pop up which together illustrate the complexity of the subject. In its simplest form, behaviour may be a series of muscle contractions, perhaps performed in clear response to a specific stimulus, such as in the case of a reflex. However, at the other extreme, we find enormously complex activities, such as birds migrating across the world, continuously assessing their direction and position with the help of various cues from stars, land- marks and geomagneticism. It may not be obvious which stimuli actually trigger the onset of this behaviour. Indeed, a bird kept in a cage in a win- dowless room with constant light will show strong attempts to escape and move towards the south at the appropriate time, without any apparent external cues at all. We would use the word behaviour for both these extremes, and for many other activities in between in complexity. It will include all types of activities that animals engage in, such as locomotion, grooming, repro- duction, caring for young, communication, etc. Behaviour may involve one individual reacting to a stimulus or a physiological change, but may also involve two individuals, each responding to the activities of the other. And why stop there? We would also call it behaviour when ani- mals in a herd or an aggregation coordinate their activities or compete for resources with one another. -
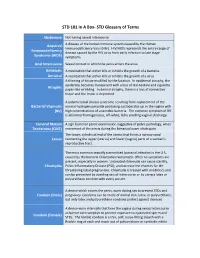
STD Glossary of Terms
STD 101 In A Box- STD Glossary of Terms Abstinence Not having sexual intercourse Acquired A disease of the human immune system caused by the Human Immunodeficiency Virus (HIV). HIV/AIDS represents the entire range of Immunodeficiency disease caused by the HIV virus from early infection to late stage Syndrome (AIDS) symptoms. Anal Intercourse Sexual contact in which the penis enters the anus. Antibiotic A medication that either kills or inhibits the growth of a bacteria. Antiviral A medication that either kills or inhibits the growth of a virus. A thinning of tissue modified by the location. In epidermal atrophy, the epidermis becomes transparent with a loss of skin texture and cigarette Atrophic paper-like wrinkling. In dermal atrophy, there is a loss of connective tissue and the lesion is depressed. A polymicrobial clinical syndrome resulting from replacement of the Bacterial Vaginosis normal hydrogen peroxide producing Lactobacillus sp. in the vagina with (BV) high concentrations of anaerobic bacteria. The common symptom of BV is abnormal homogeneous, off-white, fishy smelling vaginal discharge. Cervical Motion A sign found on pelvic examination suggestive of pelvic pathology; when Tenderness (CMT) movement of the cervix during the bimanual exam elicits pain. The lower, cylindrical end of the uterus that forms a narrow canal Cervix connecting the upper (uterus) and lower (vagina) parts of a woman's reproductive tract. The most common sexually transmitted bacterial infection in the U.S., caused by the bacteria Chlamydia trachomatis. Often no symptoms are present, especially in women. Untreated chlamydia can cause sterility, Chlamydia Pelvic Inflammatory Disease (PID), and increase the chances for life- threatening tubal pregnancies. -

Pheromones and Animal Behaviour Communication by Smell and Taste
Pheromones and Animal Behaviour Communication by Smell and Taste Tristram D. Wyatt University of Oxford published by the press syndicate of the university of cambridge The Pitt Building, Trumpington Street, Cambridge, United Kingdom cambridge university press The Edinburgh Building, Cambridge CB2 2RU, UK 40 West 20th Street, New York, NY 10011-4211, USA 477 Williamstown Road, Port Melbourne, VIC 3207, Australia Ruiz de Alarcón 13, 28014 Madrid, Spain Dock House, The Waterfront, Cape Town 8001, South Africa http://www.cambridge.org © Cambridge University Press 2003 This book is in copyright. Subject to statutory exception and to the provisions of relevant collective licensing agreements, no reproduction of any part may take place without the written permission of Cambridge University Press. First published 2003 Printed in the United Kingdom at the University Press, Cambridge Typeface Swift 9/13pt System QuarkXPress® [tb] A catalogue record for this book is available from the British Library Library of Congress Cataloguing in Publication data Wyatt, Tristram D., 1956– Pheromones and animal behaviour: communication by smell and taste / Tristram D. Wyatt. p. cm. Includes bibliographical references (p. ). ISBN 0 521 48068 X – ISBN 0 521 48526 6 (pb.) 1. Animal communication. 2. Pheromones. 3. Chemical senses. I. Title. QL776 .W93 2002 591.59 – dc21 2002024628 ISBN 0 521 48068 X hardback ISBN 0 521 48526 6 paperback The publisher has used its best endeavours to ensure that the URLs for external web sites re- ferred to in this book are correct and active at time of going to press. However, the publisher has no responsibility for the web sites and can make no guarantee that a site will remain live or that the content is or will remain appropriate. -

Description of the Chemical Senses of the Florida Manatee, Trichechus Manatus Latirostris, in Relation to Reproduction
DESCRIPTION OF THE CHEMICAL SENSES OF THE FLORIDA MANATEE, TRICHECHUS MANATUS LATIROSTRIS, IN RELATION TO REPRODUCTION By MEGHAN LEE BILLS A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2011 1 © 2011 Meghan Lee Bills 2 To my best friend and future husband, Diego Barboza: your support, patience and humor throughout this process have meant the world to me 3 ACKNOWLEDGMENTS First I would like to thank my advisors; Dr. Iskande Larkin and Dr. Don Samuelson. You showed great confidence in me with this project and allowed me to explore an area outside of your expertise and for that I thank you. I also owe thanks to my committee members all of whom have provided valuable feedback and advice; Dr. Roger Reep, Dr. David Powell and Dr. Bruce Schulte. Thank you to Patricia Lewis for her histological expertise. The Marine Mammal Pathobiology Laboratory staff especially Drs. Martine deWit and Chris Torno for sample collection. Thank you to Dr. Lisa Farina who observed the anal glands for the first time during a manatee necropsy. Thank you to Astrid Grosch for translating Dr. Vosseler‟s article from German to English. Also, thanks go to Mike Sapper, Julie Sheldon, Kelly Evans, Kelly Cuthbert, Allison Gopaul, and Delphine Merle for help with various parts of the research. I also wish to thank Noelle Elliot for the chemical analysis. Thank you to the Aquatic Animal Health Program and specifically: Patrick Thompson and Drs. Ruth Francis-Floyd, Nicole Stacy, Mike Walsh, Brian Stacy, and Jim Wellehan for their advice throughout this process.