Polyandry and Female Control: the Red Flour Beetle Tribolium Castaneum As a Case Study
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Insect Pests of Stored Grain Blog
Insect Pests of Stored Grain Insect Pest Population Potential • Insects are 1/16 to ½ inch depending on the species. • Large numbers insects in small amounts of debris. • 500 female insects • each female produces 200 offspring • 10 million insects in two generations. Adapted from the Penn State University Agronomy Guide Store Grain Insects Economic Damage • Lose up to 10% of the grain weight in a full storage bin • grain bin containing 30,000 bushels of corn valued at $3.00 per bushel would lose $9,000 • The loss does not include dockage or the cost of eliminating the insects from the grain. Adapted from the Penn State University Agronomy Guide Sampling for Bugs Looking for Bugs Docking screens can be used to separate beetles from the grain. Primary Stored Grain Feeders in NYS Weevils • Granary Weevil • Rice Weevil • Maize Weevil Beetles • Lesser Grain Borer Moths • Angoumois grain moth Weevils Have Snouts! Snout No Snout Gary Alpert, Harvard University, Bugwood.org Gary Alpert, Harvard University, Bugwood.org Maize Weevil Lesser Grain Borer Granary weevil Sitophilus granarius (L.) • polished, blackish or brown. • 3/16 of an inch long • no wings • Not in the field • longitudinal punctures- thorax • 80-300 eggs laid • One egg per grain kernel • corn, oats, barley, rye, and wheat Clemson University - USDA Cooperative Extension Slide Series , Bugwood.org Rice Weevil (Sitophilus oryzae) • 3/32 of an inch. • reddish brown to black • Small round pits-thorax • Has wings with yellow markings • Lays 80-500 eggs inside of grain • One egg per grain kernel • Start in the field • wheat, corn, oats, rye, Joseph Berger, Bugwood.org barley, sorghum, buckwheat, dried beans Maize Weevil Sitophilus zeamais • Very similar to rice weevil • slightly larger • 1/8 of an inch long • Small round pits on thorax with a mid line. -

4 Biology, Behavior, and Ecology of Insects in Processed Commodities
4 Biology, Behavior, and Ecology of Insects in Processed Commodities Rizana M. Mahroof David W. Hagstrum Most insects found in storage facilities consume Red flour beetle, Tribolium commodities, but some feed on mold growing castaneum (Herbst) on stored products. Others may be predators and parasitoids. Insects that attack relatively dry pro- Red flour beetle adults (Figure 1) are reddish brown. cessed commodities (those with about 10% or more Eggs are oblong and white. Adults show little moisture content at 15 to 42oC) can cause signifi- preference for cracks or crevices as oviposition sites. cant weight losses during storage. Insects occur in Eggshells are coated with a sticky substance that aids flour mills, rice mills, feed mills, food processing in attaching the eggs to surfaces and causes small facilities, breakfast and cereal processing facilities, particles to adhere to them (Arbogast 1991). Larvae farm storages, grain bins, grain elevators, bakeries, are yellowish white with three pair of thoracic legs. warehouses, grocery stores, pet-food stores, herbari- ums, museums, and tobacco curing barns. Economic Typically, there are six to seven larval instars, losses attributed to insects include not only weight depending on temperature and nutrition. Larvae loss of the commodity, but also monitoring and pest move away from light, living concealed in the food. management costs and effects of contamination on Full-grown larvae move to the food surface or seek product trade name reputation. shelter for pupation. Pupae are white and exarate, which means that appendages are not fused to the body. External genitalic characters on pupae can be Life Histories used to differentiate males and females (Good 1936). -

The Carcinogenic Effects of Benzoquinones Produced by the Flour Beetle
Polish Journal of Veterinary Sciences Vol. 14, No. 1 (2011), 159-164 DOI 10.2478/v10181-011-0025-8 Review The carcinogenic effects of benzoquinones produced by the flour beetle Ł.B. Lis1, T. Bakuła1, M. Baranowski1, A. Czarnewicz2 1 Department of Veterinary Prevention and Feed Hygiene Faculty of Veterinary Medicine, University of Warmia and Mazury in Olsztyn, Oczapowskiego 13, 10-718 Olsztyn, Poland 2 Veterinary Surgery HelpVet, Plonska 113A, 06-400 Ciechanów, Poland Abstract Humans and animals come into contact with various compounds in their natural environment. Most of the encountered substances are neutral, yet some may carry adverse health effects. The ingested food may be a source of harmful substances, including benzoquinones which, as shown by research results cited in this paper, demonstrate toxic, carcinogenic and enterotoxic activity. This group of compounds is inclusive of 2-methyl-1,4-benzoquinone (MBQ) and 2-ethyl-1,4-benzoquinone (EBQ), defensive secretions of the confused flour beetle (Tribolium confusum J. du V) and the red flour beetle (Tribolium castaneum Herbst). Benzoquinones have a carcinogenic effect, they are inhibi- tors of growth of various microorganisms, they produce a self-defense mechanism in threat situations and affect population aggregation. As noted by the referenced authors, the properties of ben- zoquinones have not been fully researched to this date. Key words: benzoquinones, flour beetles, quinone carcinogenicity Introduction set wider than the eyes of red flour beetle. Both spe- cies have well-formed flight wings, but only the red Flour beetles, including the confused flour beetle flour beetle flies in warm interiors or in a tropical (Tribolium confusum) and the red flour beetle climate. -

INSECTS INJURING STORED FOOD PRODUCTS in CONNECTICUT by W
CONNECTICUT AGRICULTURALEXPERIMENTSTATION NEW HAVEN, CONN. BULLETIN 195, JULY, 1917 ENTOMOLOGICAL SERIES, No. 24. -. -- INSECTS INJURING STORED FOOD PRODUCTS IN CONNECTICUT By W. E. BRITTON, Sfaft En/ontolo~q-is/. FIGUREI. Ear of corn injured by fhe European grain moth, half natural size. CONTENTS. PAGE PAGE Officers and Staff of Station ............ 2 hierlitermnean Flour hioth ............. II Insects Injuring Stored Food I'rnrlucts Jlenl Snout Moth ....................... IZ in Connecticut ........................ 3 The Grain hIoths ....................... 13 The Grain Reetles ...................... 3 r\ngolllnois Grain 310th ................ I3 Common hieal \Vorm .................. 3 European Grain Moth .................. 14 Darker Meal JVorm .................... 4 Other Insects Occasionally tittacking Cadelle .................................. 4 I'oods ................................. 14 Pea U'eevil ............................. 5 Control hlethods ........................16 Common Rean TVeevil ................. 6 Temperature ............................ r6 Four-Spotted Rean TVeevil. ............. 7 Hent ....................................16 Drug Store Ileetle ...................... 7 Cold .................................... 17 Confused and Rust-Red lzlour Heetles. 8 Air-Slaked Lime ........................ 17 Saw-toothed Grain Heetle .............. 8 Pest-proof Packages .................. 18 Granary Weevil ....................... 9 Fumigation ............................. rq Rice M'eevil ............................ 9 Carbon Disulphide -

01 6050 Houseflies (Page 1)
Aventis Environmental Science Confused flour beetle Flour Beetles 3.5mm long Description Order: Coleoptera (‘sheath-wings’) Characteristics: Forewings hard and leathery, meeting along mid- line of dorsal surface; hindwings membranous, sometimes lacking; biting mouthparts; well developed thorax; complete metamorphosis with egg, larval, pupal and adult stages. Family: Tenebrionidae Usually 11-segmented antennae; front and middle tarsi are 5-segmented and hind tarsi 4-segmented. Species characteristics: Confused Flour Beetle (Tribolium confusum) Adult elongate, 2.6-4.4mm in length; colour, reddish-brown; antennae with loose, indistinct, 5 or 6 segmented club which may be lacking; slight ridge evident above each eye. Distribution Life-Cycle Rust-Red Flour Beetle Flour beetles are cosmopolitan. Two of the most Up to 450 eggs are laid singly at a rate of 2-10 (Tribolium castaneum) Adult elongate, common species are the Confused flour beetle, per day, depending on temperature. The white 2.3-4.4mm in length; colour, reddish-brown; which is commonly encountered in bakeries and eggs are sticky and rapidly become coated with antennae with distinct, 3-segmented club at tip; flour mills, and the Rust-red flour beetle. Both food particles and other debris. At 22-27 °C they no ridge present above each eye. are serious pests of cereal products, including hatch in 6-14 days. The larvae are white tinged grain, flour, porridge oats and rice bran. with yellow and pass through 5-11 moults Dark Flour Beetle before reaching a full-grown length of 5mm. (Tribolium destructor) Adult elongate, Other products which may be attacked are oil This process takes 3-9 weeks. -
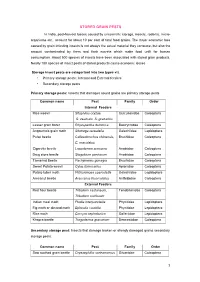
Stored Grain Pests
STORED GRAIN PESTS In India, post-harvest losses caused by unscientific storage, insects, rodents, micro- organisms etc., account for about 10 per cent of total food grains. The major economic loss caused by grain infesting insects is not always the actual material they consume, but also the amount contaminated by them and their excreta which make food unfit for human consumption. About 500 species of insects have been associated with stored grain products. Nearly 100 species of insect pests of stored products cause economic losses Storage insect pests are categorized into two types viz. • Primary storage pests : Internal and External feeders • Secondary storage pests Primary storage pests: Insects that damages sound grains are primary storage pests Common name Pest Family Order Internal Feeders Rice weevil Sitophilus oryzae, Curculionidae Coleoptera S. zeamais, S. granarius Lesser grain borer Rhyzopertha dominica Bostrychidae Coleoptera Angoumois grain moth Sitotroga cerealella Gelechiidae Lepidoptera Pulse beetle Callosobruchus chinensis, Bruchidae Coleoptera C. maculatus Cigarette beetle Lasioderma sericorne Anobiidae Coleoptera Drug store beetle Stegobium paniceum Anobiidae Coleoptera Tamarind Beetle Pachymeres gonagra Bruchidae Coleoptera Sweet Potato weevil Cylas formicarius Apionidae Coleoptera Potato tuber moth Phthorimoea operculella Gelechiidae Lepidoptera Arecanut beetle Araecerus fasciculatus Anthribidae Coleoptera External Feeders Red flour beetle Tribolium castaneum, Tenebrionidae Coleoptera Tribolium confusum Indian meal -

Confused Flour Beetle Tribolium
Confused Flour Beetle Tribolium Species: confusum Genus: Tribolium Family: Tenebrionidae Order: Coleoptera Class: Insecta Phylum: Arthropoda Kingdom: Animalia Conditions for Customer Ownership We hold permits allowing us to transport these organisms. To access permit conditions, click here. Never purchase living specimens without having a disposition strategy in place. There are currently no USDA permits required for this organism. In order to protect our environment, never release a live laboratory organism into the wild. Primary Hazard Considerations • Always wash your hands after handling these beetles. • There are no health risks from the Confused Flour beetle (even if ingested!). They will not bite. • They can become pests if released in households. They infest dry, stored foods such as cereal, grain, beans, dried fruit, nuts, flour, and even chocolate. Take care not to let any escape. The best way to prevent an infestation is to keep storage areas clean and free from any food debris. • Avoid breathing residual dust. Availability • No seasonality. Confused Flour beetles are bred in our labs, so are generally available year around. • The larvae, pupae, and adults can be found throughout the substrate. • Beetles will arrive packed in a 4 oz. jar in flour with a damp paper towel. We over-pack each order of beetles. It is normal to have some deceased beetles in the container. You will receive at least the quantity of live beetles stated on the container. Healthy larvae 1 and adults are very active, but they do not fly. Adults are about ⁄8 inch long and reddish brown. They can live in the container they are shipped in for about 7–10 days before needing to be transferred to a habitat. -

Tribolium and Friends Meeting EURO EVO DEVO June 25Th-26Th Galway 2018
Abstract Booklet Tribolium and Friends Meeting EURO EVO DEVO June 25th-26th Galway 2018 i Table of Contents Meeting Schedule ................................................................................................................. iii Session 1 ............................................................................................................................... 1 Ariel Chipman. Oncopeltus, Tribolium, Drosophila – a three-taxon problem for understanding the evolution of segmentation in insects ......................................................... 1 Martin Klingler. Double abdomen in a short germ insect: Zygotic control of axis formation revealed in the beetle Tribolium castaneum ........................................................................... 2 Rahul Sharma. Segmentation in Arthropods: a study investigating regulatory relationship between gap genes and pair-rule genes in the beetle Tribolium castaneum .......................... 3 Matt Benton. A revised understanding of Tribolium morphogenesis further reconciles short and long germ development ................................................................................................... 4 Session 2 ................................................................................................................................ 5 Allistair McGregor. Spider segmentation gets its Sox on ................................................... 5 Olivia Tidswell. Using Hybridisation Chain Reaction to study Hox gene regulation in Tribolium .............................................................................................................................. -

Genome of the Small Hive Beetle (Aethina Tumida, 2004;32(5):1792–7
GigaScience, 7, 2018, 1–16 doi: 10.1093/gigascience/giy138 Advance Access Publication Date: 7 December 2018 Research RESEARCH Genome of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae), a worldwide parasite of social bee colonies, provides insights into detoxification and herbivory Jay D. Evans 1,*, Duane McKenna2,ErinScully3, Steven C. Cook1, Benjamin Dainat, 4, Noble Egekwu1, Nathaniel Grubbs5, Dawn Lopez1, Marce´ D. Lorenzen5,StevenM.Reyna5, Frank D. Rinkevich6, Peter Neumann7 and Qiang Huang7,8,* 1USDA-ARS, Bee Research Laboratory, BARC-East Building 306, Beltsville, Maryland 20705, USA, 2Department of Biological Sciences, University of Memphis, 3700 Walker Ave., Memphis, TN 38152, USA, 3USDA-ARS, Center for Grain and Animal Health, Stored Product Insect and Engineering Research Unit, Manhattan, KS 66502, USA, 4Agroscope, Swiss Bee Research Center, CH-3003 Bern, Switzerland, 5Department of Entomology and Plant Pathology, North Carolina State University, 1566 Thomas Hall, Raleigh, NC 27695, USA, 6USDA, Honey Bee Breeding, Genetics and Physiology Laboratory, 1157 Ben Hur Road, Baton Rouge, LA 70820, USA, 7Institute of Bee Health, Vetsuisse Faculty, University of Bern, Schwarzenburgstrasse 161, CH-3097, Liebefeld, Switzerland and 8Honey Bee Research Institute, Jiangxi Agricultural University, Zhimin Avenue 1101, 330045 Nanchang, China | downloaded: 10.10.2021 ∗Correspondence address. Jay D. Evans, USDA-ARS, Bee Research Laboratory, BARC-East Building 306, Beltsville, Maryland 20705, USA. E-mail: [email protected] http://orcid.org/0000-0002-0036-4651; Qiang Huang, E-mail: [email protected] Abstract Background: The small hive beetle (Aethina tumida; ATUMI) is an invasive parasite of bee colonies. ATUMI feeds on both fruits and bee nest products, facilitating its spread and increasing its impact on honey bees and other pollinators. -

A. Primary Storage Pests
STORED GRAIN PESTS: In India, post-harvest losses caused by unscientific storage, insects, rodents, microorganisms etc., account for about 10 per cent of total food grains. The major economic loss caused by grain infesting insects is not always the actual material they consume, but also the amount contaminated by them and their excreta which make food unfit for human consumption. About 500 species of insects have been associated with stored grain products. Nearly 100 species of insect pests of stored products cause economic losses. Storage insect pests are categorized into two types viz. • Primary storage pests : Internal and External feeders • Secondary storage pests Primary storage pests: Insects that damages sound grains are primary storage pests. Some of examples are: Rice weevil Sitophilus oryzae, Lesser grain borer: Rhyzopertha dominica Bostrychidae Coleoptera Pulse beetle: Callosobruchus chinensis, Red flour beetle: Tribolium castaneum, Tribolium confusum Rice moth: Corcyra cephalonica Khapra beetle: Trogoderma granarium Dermestidae Coleoptera Secondary storage pest: Insects that damage broken or already damaged grains secondary storage pests. Examples: Saw toothed grain beetle: Oryzaephillis surinamensis Silvanidae: Coleoptera Long headed flour beetle: Latheticus oryzae Tenebrionidae Coleoptera Flat grain beetle: Cryptolestus minutas, Cucujidae Coleoptera Grain lice: Liposcelis divinitorius Liposcelidae Psocoptera Grain mite : Acarus siro Acari A. Primary storage pests: • Rice weevil: Sitophilus oryzae Classification: Phylum: Arthropoda Class: Insecta Family : Cruculionidae Genus: Sitophilus Species: oryzae Diagonostic features: • Adults are reddish brown having a length of 3 mm with a long, slender, pointed head. • Full grown larva is 5 mm in length and plumpy, fleshy, legless creature. Its elytra bear four light reddish or yellowish spots. • The rice weevil breeds from April to October and hibernates in winter as an adult inside cracks and crevices or under wheat bags in the godowns. -

Beetles in Flour, Grains, and Other Stored Foods
INSECT DIAGNOSTIC LABORATORY IDL Cornell University, Dept. of Entomology, 2144 Comstock Hall, Ithaca NY 14853-2601 Beetles in Flour, Grains, and Other Stored Foods Confused flour beetle. (Red flour beetle looks similar.) Yellow mealworm Photo by Gary Alpert, larva (above) Harvard University. and adult (to right). Photos from Clemson University, USDA Cooperative Extension Slide Series, Bugwood.org Injury Beetles of several species infest packages of whole grain and grain products. The infestation may begin at the time of manufacture or processing, in the warehouses of food distributors, in transit, on the grocers' shelves, or in the home. Most food processors and handlers make every effort to avoid insect infestations, but occasionally the efforts fail. Infestations are usually discovered when an infested package is opened for use, or when small brown beetles are found in the kitchen near containers of stored grain products. A wide variety of foods may be infested, including flour, cereal, dried fruits, dehydrated vegetables, shelled nuts, chocolate, spices, candies, pet foods, and bird seed. Eggs, larvae, pupae, and adults of the beetles may occur in infested foods. Description Two dozen or more different species of insects may occasionally infest grain and grain products used in homes, but four species are much more frequent than the others. Three of these are minute insects, and the fourth is moderate in size. The confused flour beetle (Tribolium confusum) is perhaps the most common. It is about 1/7 inch in length, an elongated, dark brown, hard-shelled beetle. Nearly as common is the sawtoothed grain beetle (Oryzaephilus surinamensis), which is slightly shorter and more slender. -

Red Flour Beetle, Tribolium Castaneum
Rationale to sequence the genome of the red flour beetle, Tribolium castaneum Susan Brown1, Robin Denell1, Richard Beeman2 and Richard Gibbs3 1Division of Biology, Ackert Hall, Kansas State University, Manhattan, KS 2 USDA-ARS-GMPRC, Manhattan, KS 3 HGSC, Baylor College of Medicine, Houston, TX Overview: In a world of worthy candidates, there are several compelling reasons to sequence the genome of the red flour beetle Tribolium castaneum. First and foremost, Tribolium is one of the most sophisticated genetic model organisms among all higher eukaryotes. Among arthropods, only Drosophila offers greater power and flexibility of genetic manipulation. Second, the Tribolium genome sequence will provide an informative link when direct comparisons between human and fruit fly sequences are unproductive. Third, as a member of the most primitive order of holometabolous insects, the Coleoptera, it is in a key phylogenetic position to inform us about the genetic innovations that accompanied the evolution of higher forms with more complex development. Fourth, Coleoptera is the largest and most species diverse of all eukaryotic orders and Tribolium offers the only genetic model for this profusion of medically and economically important species. Analysis of the Tribolium genome will facilitate the discovery of new pharmaceuticals and antibiotics. In addition, it will lead to a better understanding of resistance mechanisms and improved specificity and efficacy of insecticides and biological agents to control crop pests and disease vectors. Finally, many genetic and genomic tools are already available for Tribolium. Both forward and reverse genetic approaches are available to facilitate functional genetic analysis. A molecular recombination map and a physical map based on a BAC library of ~20 x genome coverage are now being completed, and will furnish the necessary foundation upon which to assemble the genomic sequence, which at ~200 Mbs is only slightly larger than that of Drosophila.