1 a Naturalist on the Edge of Evolution: Why Thomas Nagel Should Embrace ID
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Argumentation and Fallacies in Creationist Writings Against Evolutionary Theory Petteri Nieminen1,2* and Anne-Mari Mustonen1
Nieminen and Mustonen Evolution: Education and Outreach 2014, 7:11 http://www.evolution-outreach.com/content/7/1/11 RESEARCH ARTICLE Open Access Argumentation and fallacies in creationist writings against evolutionary theory Petteri Nieminen1,2* and Anne-Mari Mustonen1 Abstract Background: The creationist–evolutionist conflict is perhaps the most significant example of a debate about a well-supported scientific theory not readily accepted by the public. Methods: We analyzed creationist texts according to type (young earth creationism, old earth creationism or intelligent design) and context (with or without discussion of “scientific” data). Results: The analysis revealed numerous fallacies including the direct ad hominem—portraying evolutionists as racists, unreliable or gullible—and the indirect ad hominem, where evolutionists are accused of breaking the rules of debate that they themselves have dictated. Poisoning the well fallacy stated that evolutionists would not consider supernatural explanations in any situation due to their pre-existing refusal of theism. Appeals to consequences and guilt by association linked evolutionary theory to atrocities, and slippery slopes to abortion, euthanasia and genocide. False dilemmas, hasty generalizations and straw man fallacies were also common. The prevalence of these fallacies was equal in young earth creationism and intelligent design/old earth creationism. The direct and indirect ad hominem were also prevalent in pro-evolutionary texts. Conclusions: While the fallacious arguments are irrelevant when discussing evolutionary theory from the scientific point of view, they can be effective for the reception of creationist claims, especially if the audience has biases. Thus, the recognition of these fallacies and their dismissal as irrelevant should be accompanied by attempts to avoid counter-fallacies and by the recognition of the context, in which the fallacies are presented. -
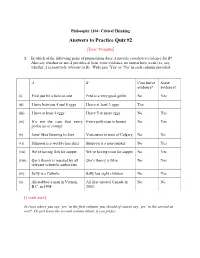
Answers to Practice Quiz #2
Philosophy 1104: Critical Thinking Answers to Practice Quiz #2 [Total: 50 marks] 1. In which of the following pairs of propositions does A provide conclusive evidence for B? Also say whether or not A provides at least some evidence, no matter how weak (i.e. say whether A is positively relevant to B). Write just „Yes‟ or „No‟ in each column provided. A B Conclusive Some evidence? evidence? (i) Fred just hit a hole-in-one Fred is a very good golfer No Yes (ii) I have between 4 and 6 eggs I have at least 3 eggs Yes (iii) I have at least 4 eggs I have 5 or more eggs No Yes (iv) It‟s not the case that every Every politician is honest No Yes politician is corrupt. (v) Janet likes listening to Jazz Vancouver is west of Calgary No No (vi) Simpson is a world-class skier Simpson is a non-smoker No Yes (vii) We‟re having fish for supper We‟re having trout for supper No Yes (viii) Qin‟s theory is rejected by all Qin‟s theory is false No Yes relevant scientific authorities (ix) Sally is a Catholic Sally has eight children No Yes (x) Ali stabbed a man in Vernon, Ali first entered Canada in No No B.C. in 1998. 2002. [1 mark each] In rows where you say ‘yes’ in the first column, you should of course say ‘yes’ in the second as well! Or just leave the second column blank, if you prefer. 2. Each of the following definitions is flawed in some way (each in just one way, I think, or at least one main one). -

Probability, Statistics, Evolution, and Intelligent Design
Probability, Statistics, Evolution, and Intelligent Design Peter Olofsson n the last decades, arguments against Darwinian evolution An ID Hypothesis Testing Challenge to Evolution have become increasingly sophisticated, replacing Cre- In his book The Design Inference, William Dembski introduces ationism by Intelligent Design (ID) and the book of Genesis I the “explanatory filter” as a device to rule out chance expla- by biochemistry and mathematics. As arguments claiming to nations and infer design of observed phenomena. The filter be based in probability and statistics are being used to justify also appears in his book No Free Lunch, where the description the anti-evolution stance, it may be of interest to readers of differs slightly. In essence, the filter is a variation on statistical CHANCE to investigate methods and claims of ID theorists. hypothesis testing with the main difference being that it aims at ruling out chance altogether, rather than just a specified Probability, Statistics, and Evolution null hypothesis. Once all chance explanations have been ruled The theory of evolution states in part that traits of organisms out, ‘design’ is inferred. Thus, in this context, design is merely are passed on to successive generations through genetic mate- viewed as the complement of chance. rial and that modifications in genetic material cause changes To illustrate the filter, Dembski uses the example of in appearance, ability, function, and survival of organisms. Nicholas Caputo, a New Jersey Democrat who was in charge Genetic changes that are advantageous to successful repro- of putting together the ballots in his county. Names were to be duction over time dominate and new species evolve. -

Irreducible Complexity (IC) Has Played a Pivotal Role in the Resurgence of the Creationist Movement Over the Past Two Decades
Irreducible incoherence and Intelligent Design: a look into the conceptual toolbox of a pseudoscience Abstract The concept of Irreducible Complexity (IC) has played a pivotal role in the resurgence of the creationist movement over the past two decades. Evolutionary biologists and philosophers have unambiguously rejected the purported demonstration of “intelligent design” in nature, but there have been several, apparently contradictory, lines of criticism. We argue that this is in fact due to Michael Behe’s own incoherent definition and use of IC. This paper offers an analysis of several equivocations inherent in the concept of Irreducible Complexity and discusses the way in which advocates of the Intelligent Design Creationism (IDC) have conveniently turned IC into a moving target. An analysis of these rhetorical strategies helps us to understand why IC has gained such prominence in the IDC movement, and why, despite its complete lack of scientific merits, it has even convinced some knowledgeable persons of the impending demise of evolutionary theory. Introduction UNTIL ITS DRAMATIC legal defeat in the Kitzmiller v. Dover case, Intelligent Design Creationism (IDC) had been one of the most successful pseudosciences of the past two decades, at least when measured in terms of cultural influence. It is interesting to explore the way this species of creationism had achieved this success, notwithstanding its periodic strategic setbacks as well as its complete lack of scientific merits. Of course, a full explanation would include religious, socio-political, and historical reasons; instead, in this paper, we will take a closer look at the conceptual toolbox and rhetorical strategies of the ID creationist. -

Clarity and Confusion
Book Reviews Clarity and confusion to overcome various antibiotics, such A review of as chloroquine, and humans have The Edge of Evolution: mutated to generate some measure of The Search for the resistance to malaria (e.g., sickle cell, Limits of Darwinism thalassemia). by Michael J. Behe Behe shows that all the cases of Free Press, New York, adaptation, in both Plasmodium and NY, 2007 humans, are due to breaking things, not creating new complex features. For example, chloroquine resistance Don Batten in Plasmodium is due to a fault in a transport protein that moves the poison his new book by Michael Behe, a into the organism’s vacuole. Behe Tfollow-up to Darwin’s Black Box likens the struggle to trench warfare, (DBB), has created somewhat of a where the defending forces will destroy storm amongst the faithful defenders their own bridge, or blow up a road, of Darwin such as Richard Dawkins, to impede the enemy’s advance. It is Jerry Coyne and Kenneth Miller. They not really an arms race, because in an arms race the opposing forces invent came out with all guns blazing to try overcoming other anti-malarials has to destroy the credibility of this book, new weapons, but the natural processes (‘evolution’) operating in Plasmodium only needed one mutation for each presumably hoping that their dismissive one. This seems like a reasonable vitriol would cause potential readers to and humans have not invented new weapons. explanation for the relative resilience skip reading it. Dawkins claimed that of chloroquine over time compared to Behe had completely departed from the Chloroquine resistance—a other anti-malarials. -

The College Student's Back to School Guide to Intelligent Design
Revised November, 2014 Part I: Letter of Introduction: Why this Student’s Guide? Part II: What is Intelligent Design? Part III: Answers to Your Professors’ 10 Most Common Misinformed Objections to Intelligent Design (1) Intelligent Design is Not Science (2) Intelligent Design is just a Negative Argument against Evolution (3) Intelligent Design Rejects All of Evolutionary Biology (4) Intelligent Design was Banned from Schools by the U.S. Supreme Court (5) Intelligent Design is Just Politics (6) Intelligent Design is a Science Stopper (7) Intelligent Design is “Creationism” and Based on Religion (8) Intelligent Design is Religiously Motivated (9) Intelligent Design Proponents Don’t Conduct or Publish Scientific Research (10) Intelligent Design is Refuted by the Overwhelming Evidence for Neo-Darwinian Evolution Part IV: Information About the Discovery Institute’s Summer Seminars on Intelligent Design COPYRIGHT © DISCOVERY INSTITUTE, 2014 — WWW.INTELLIGENTDESIGN.ORG PERMISSION GRANTED TO COPY AND DISTRIBUTE FOR NONPROFIT EDUCATIONAL PURPOSES. 2 Part I: Letter of Introduction: Why this Student’s Guide? Welcome to College, Goodbye to Intelligent Design? The famous Pink Floyd song that laments, “We don’t need no education / We don’t need no thought control,” is not just the rant of a rebellious mind; it is also a commentary on the failure of education to teach students how to think critically and evaluate both sides of controversial issues. Few scientists understood the importance of critical thinking better than Charles Darwin. When he first proposed his theory of evolution in Origin of Species in 1859, Darwin faced intense intellectual opposition from both the scientific community and the culture of his day. -

The Great Mutator Lution of Many of Which We Do Not Yet Fully Understand
living species, each of which had a slightly more advanced version of an eye. Behe’s Jerry A. Coyne novelty was to extend this argument to complex biochemical pathways, the evo- THE GREAT MUTATOR lution of many of which we do not yet fully understand. It was in the complexity of metabolism, blood clotting, and immu- THE EDGE OF EVOLUTION: this disclaimer is perfectly understand- nology that Behe claimed to have found THE SEARCH FOR THE LIMITS able. For in this department resides Mi- the hand of the Great Designer. OF DARWINISM chael Behe—that rara avis, a genuine The reviews of Darwin’s Black Box in By Michael J. Behe biologist who is also an advocate of “in- the scientific community were uniformly (Free Press, 320 pp., $28) telligent design.” And Lehigh University negative, for two reasons. First, we do does not wish to lose prospective stu- understand something about how these I. dents who bridle at the thought of study- pathways might have evolved in stepwise rowsing the websites of dif- ing miracles in their science courses. fashion, though we are as yet admittedly ferent colleges, a prospective Intelligent design, or ID, is a modern ignorant of many details. (It is harder to biology student finds an un- form of creationism cleverly constructed reconstruct the evolution of biochemi- usual statement on the page of to circumvent the many court decisions cal pathways than the evolution of or- the Department of Biological that have banned, on First Amendment ganisms themselves, because, unlike Sciences at Lehigh University. It begins: grounds, the teaching of religious views organisms, these pathways do not fos- B in the science classroom. -

Carroll on Behe-Intelligent Design
BOOKS ET AL. EVOLUTION proteins evolve and how proteins interact, and he completely ignores a huge amount of God as Genetic Engineer experimental data that directly contradicts his faulty premises. Unfortunately, these errors Sean B. Carroll are of a technical nature and will be difficult for lay readers, and even some scientists “The Lord hath delivered him into mine such as vertebrate classes). He also accepts (as (those unfamiliar with molecular biology and hands.” he has before) the 4.5-billion-year age of evolutionary genetics), to detect. Some people Earth and that we share a common ancestor will be hoodwinked. My goal here is to point hose are the words that Thomas Hux- with chimpanzees. That certainly won’t go out the critical flaws in Behe’s key arguments ley, Darwin’s confidant and staunchest over well in some camps. and to guide readers toward ally, purportedly murmured to a col- Behe also explores some ex- some references that illus- T The Edge of Evolution league as he rose to turn Bishop Samuel amples of Darwinian evolution trate why what he alleges to Wilberforce’s own words to his advantage and at the molecular level, including The Search for the be beyond the limits of rebut the bishop’s critique of Darwin’s theory an extensive treatment of the Limits of Darwinism Darwinian evolution falls at their legendary 1860 Oxford debate. They evolutionary “trench warfare” by Michael J. Behe well within its demonstrated are also the first words that popped into my fought between humans and Free Press, New York, 2007. -

Falling Over the Edge Claims That an Intelligent Designer Is Needed to Explain Evolution of Complex Systems Are Deeply Flawed
Vol 447|28 June 2007 BOOKS & ARTS Falling over the edge Claims that an intelligent designer is needed to explain evolution of complex systems are deeply flawed. The Edge of Evolution: The Search for the Limits of Darwinism by Michael Behe Free Press: 2007. 336 pp. $28 Kenneth R. Miller Michael Behe’s new book, The Edge of Evolu- tion, is an attempt to give the intelligent-design movement a bit of badly needed scientific sup- port. After a spectacular setback in the 2005 Dover, Pennsylvania, intelligent-design trial (Nature 439, 6–7; 2006), and the 2006 elec- toral losses in Ohio and Kansas, the movement could use some help — and Behe is eager to provide it. Knowing how easy it is to demonstrate the workings of evolution in the development of drug resistance in viruses, bacteria and pro- tozoan parasites, Behe concedes the point that evolution works very well at this level. His case study, repeated almost to the point of tedium, is malaria. Resistance to drugs such as chloroquine has indeed arisen within the parasite population, and so has genetic resist- ance to the parasite in humans. But if the inter- Far from finding a clean edge to evolution, Behe’s poorly designed arguments crumble away. species genetic warfare between Homo sapiens and Plasmodium is actually a prime example of badly designed argument collapses under its these two exact mutations occurring simultan- evolution, how can it then be used to make the own weight. eously at precisely the same position in exactly case for ‘design’? Behe cites the malaria literature to note the same gene in a single individual. -

Tensions in Intelligent Design's Critique of Theistic Evolutionism
Tensions in Intelligent Design’s Critique of Theistic Evolutionism _________________________________________________________ Erkki Vesa Rope Kojonen Abstract “Intelligent Design” (ID) is a contemporary intellectual movement arguing that there is scientific evidence for the existence of some sort of creator. Its proponents see ID as a scientific research program and as a way of building a bridge between science and theology, while many critics see it merely as a repackaged form of religiously motivated creationism: both bad science and bad theology. In this article, I offer a close reading of the ID movement’s critique of theistic evolutionism (TE) and argue that this critique is ultimately in tension with the movement’s broader thought. Introduction The Intelligent Design movement argues that the order of nature provides scientific evidence of the existence of an “intelligent designer”, typically believed to be the God of the Bible. Intelligent Design is a counter-reaction to the atheistic evolutionism of thinkers like Richard Dawkins (1991), who argue that evolutionary theory shows the unnecessity of a Creator. However, from its beginnings ID has also critiqued theistic evolutionism, which accepts both the doctrine of creation and the mainstream scientific theory of Darwinian evolution. This article has a twofold purpose. First, I aim to increase understanding of Intelligent Design and its central differences from theistic evolutionism. Second, I will argue that ID’s critique of theistic evolutionism is ultimately in tension with the movement’s broader thought. Thus, this critique should be abandoned or substantially modified even by proponents of ID. Theistic evolutionism and ID have some similarity, because both oppose atheistic evolutionism. -

PE1530/A: Centre for Intelligent Design UK
PE1531/A Response to Petition 1530 re Creationism in Schools Submission from the Centre for Intelligent Design UK October 2014 On behalf of the Centre for Intelligent Design, we wish to raise serious concerns about Public Petition 1530 submitted by Spencer Fildes on behalf of the Scottish Secular Society. We request that the Petitions Committee consider the position outlined in this submission. Note: Section 1 contains the main points we wish the Petitions Committee to consider. Sections 2-7 contain further explanatory and background material. Some of the points made in the summary require elaboration in the interests of clarity. 1.0 Summary 1.1 In our view, this petition is significantly misguided and would be an unsafe basis for making any changes to Government guidance on science education in the area of origins. It is highly inaccurate in its terminology and in its view of the scientific method and of educational pedagogy. 1.2 It is particularly disturbing that the Petition does not recognise the difference between ‘creationism’ and ‘intelligent design’. Failure to make this distinction leads to a distorted and inaccurate analysis of the how the study of origins should be handled in schools. Intelligent Design, properly understood, is a minimal commitment to intelligent causation in nature and is a legitimate inference from scientific data. It is not a religious position like ‘creationism’ and should not therefore be discounted in science education. It is also inaccurate to say that intelligent design is ‘an alternative to evolution’. 1.3 The Petition also contains a fundamental misunderstanding about the nature of scientific controversy. -

The Edge of Evolution the Search for the Limits of Darwinism By: Michael J
╝ The Edge of Evolution The Search for The Limits of Darwinism By: Michael J. Behe ]1[ خ نبذة ُمترصة عن الكتاب: كتاب آخر هام جد ًا للعامل املشهور جد ًا املُتخ ِّصص يف جمال الكيمياء احليوية: »مايكل بيهي«. كتاب »بيهي« اﻷول، والذي كان ب ُعنوان »صندوق داروين اﻷسود«، كان يتك َّلم عن فكرة »ال َّتعقيد غري القابل لﻻختزا ل«، وكان ينتقد هبذه الفكرة إمكانية أو ُقدرة نظرية ال َّتط ُّور ال َّداروينية عىل أن ُتق ِّدم تفسرياً لنشأة احلياة، ومل يتع َّرض »بيهي« للمفهوم ال َّت قليدي لنظرية ال َّتطور ال َّداروينية، فيام ُخي ّص تفسري ال َّتن ُّوع احلاصل بني الكائنات احل َّية. يف هذا الكتاب، الذي هو ب ُعنوان »حا َّفة ال َّتط ُّور« أو »ح ّد ال َّت ط ُّور«، ُيناقش »بيهي« ُقدرة نظرية ال َّتط ُّور ال َّداروينية عىل تفسري ال َّتن ُّ عو بني الكائنات احل َّي ،ة واهلدف من الكتاب هو بيان أن ُعنرص ّي الن َّظرية: الطفرات العشوائية واﻻنتقاء الطبيعي، غري قادران ب ُم فردمها عىل تفسري أشكال احلياة املُختلفة عىل اﻷرض! لعل هذا الكتاب هو اﻷهم فيام ُخيص نقد نظرية ال َّتط ُّور ال َّداروينية، وﻻ أريد أن أطيل الكﻻم كثرياً عن الكتاب، فهو – ككتابه اﻷول – ميلء باملعلومات العلمية والتفاصيل الكثرية الصعبة واملُم َّلة، ولك َّن أه ّم ما يف الكتاب هو تقريرا ت »بيهي« واستنتاجاته العلمية املبنية عىل ام قام برسده من دراسات وأبحاث. مع انتهائي من هذا الكتاب، بدأت أفهم تص ُّور »بيهي« لنظرية ال َّتط ُّور ال َّداروينية، وهذه لفتة ها َّم ة، نافعة فيام ُخي ّص طريقة ال َّتعامل مع النظرية العلمية: بيهي يستخدم تعبري »ال َّتط ُّور ال َّد ارويني« لﻹشارة إىل تغ ُّري ناتج عن عشوائية عمياء، ويقوم باستخدام كلمة »تط ُّور« ُمفردة، بمعنى عام مقبول لدى اجلميع، وهو اﻹيامن بأ َّن الكائنات احل َّية تتغ َّري مع الوقت، وبينها وبني الكائنات احل َّية اﻷخرى اختﻻفات.