Irreducible Complexity (IC) Has Played a Pivotal Role in the Resurgence of the Creationist Movement Over the Past Two Decades
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Understanding the Intelligent Design Creationist Movement: Its True Nature and Goals
UNDERSTANDING THE INTELLIGENT DESIGN CREATIONIST MOVEMENT: ITS TRUE NATURE AND GOALS A POSITION PAPER FROM THE CENTER FOR INQUIRY OFFICE OF PUBLIC POLICY AUTHOR: BARBARA FORREST, Ph.D. Reviewing Committee: Paul Kurtz, Ph.D.; Austin Dacey, Ph.D.; Stuart D. Jordan, Ph.D.; Ronald A. Lindsay, J. D., Ph.D.; John Shook, Ph.D.; Toni Van Pelt DATED: MAY 2007 ( AMENDED JULY 2007) Copyright © 2007 Center for Inquiry, Inc. Permission is granted for this material to be shared for noncommercial, educational purposes, provided that this notice appears on the reproduced materials, the full authoritative version is retained, and copies are not altered. To disseminate otherwise or to republish requires written permission from the Center for Inquiry, Inc. Table of Contents Section I. Introduction: What is at stake in the dispute over intelligent design?.................. 1 Section II. What is the intelligent design creationist movement? ........................................ 2 Section III. The historical and legal background of intelligent design creationism ................ 6 Epperson v. Arkansas (1968) ............................................................................ 6 McLean v. Arkansas (1982) .............................................................................. 6 Edwards v. Aguillard (1987) ............................................................................. 7 Section IV. The ID movement’s aims and strategy .............................................................. 9 The “Wedge Strategy” ..................................................................................... -

Intelligent Design, Abiogenesis, and Learning from History: Dennis R
Author Exchange Intelligent Design, Abiogenesis, and Learning from History: Dennis R. Venema A Reply to Meyer Dennis R. Venema Weizsäcker’s book The World View of Physics is still keeping me very busy. It has again brought home to me quite clearly how wrong it is to use God as a stop-gap for the incompleteness of our knowledge. If in fact the frontiers of knowledge are being pushed back (and that is bound to be the case), then God is being pushed back with them, and is therefore continually in retreat. We are to find God in what we know, not in what we don’t know; God wants us to realize his presence, not in unsolved problems but in those that are solved. Dietrich Bonhoeffer1 am thankful for this opportunity to nature, is the result of intelligence. More- reply to Stephen Meyer’s criticisms over, this assertion is proffered as the I 2 of my review of his book Signature logical basis for inferring design for the in the Cell (hereafter Signature). Meyer’s origin of biological information: if infor- critiques of my review fall into two gen- mation only ever arises from intelli- eral categories. First, he claims I mistook gence, then the mere presence of Signature for an argument against bio- information demonstrates design. A few logical evolution, rendering several of examples from Signature make the point my arguments superfluous. Secondly, easily: Meyer asserts that I have failed to refute … historical scientists can show that his thesis by not providing a “causally a presently acting cause must have adequate alternative explanation” for the been present in the past because the origin of life in that the few relevant cri- proposed candidate is the only known tiques I do provide are “deeply flawed.” cause of the effect in question. -

Argumentation and Fallacies in Creationist Writings Against Evolutionary Theory Petteri Nieminen1,2* and Anne-Mari Mustonen1
Nieminen and Mustonen Evolution: Education and Outreach 2014, 7:11 http://www.evolution-outreach.com/content/7/1/11 RESEARCH ARTICLE Open Access Argumentation and fallacies in creationist writings against evolutionary theory Petteri Nieminen1,2* and Anne-Mari Mustonen1 Abstract Background: The creationist–evolutionist conflict is perhaps the most significant example of a debate about a well-supported scientific theory not readily accepted by the public. Methods: We analyzed creationist texts according to type (young earth creationism, old earth creationism or intelligent design) and context (with or without discussion of “scientific” data). Results: The analysis revealed numerous fallacies including the direct ad hominem—portraying evolutionists as racists, unreliable or gullible—and the indirect ad hominem, where evolutionists are accused of breaking the rules of debate that they themselves have dictated. Poisoning the well fallacy stated that evolutionists would not consider supernatural explanations in any situation due to their pre-existing refusal of theism. Appeals to consequences and guilt by association linked evolutionary theory to atrocities, and slippery slopes to abortion, euthanasia and genocide. False dilemmas, hasty generalizations and straw man fallacies were also common. The prevalence of these fallacies was equal in young earth creationism and intelligent design/old earth creationism. The direct and indirect ad hominem were also prevalent in pro-evolutionary texts. Conclusions: While the fallacious arguments are irrelevant when discussing evolutionary theory from the scientific point of view, they can be effective for the reception of creationist claims, especially if the audience has biases. Thus, the recognition of these fallacies and their dismissal as irrelevant should be accompanied by attempts to avoid counter-fallacies and by the recognition of the context, in which the fallacies are presented. -
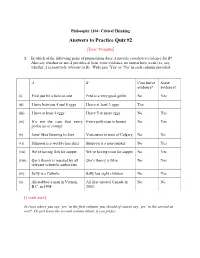
Answers to Practice Quiz #2
Philosophy 1104: Critical Thinking Answers to Practice Quiz #2 [Total: 50 marks] 1. In which of the following pairs of propositions does A provide conclusive evidence for B? Also say whether or not A provides at least some evidence, no matter how weak (i.e. say whether A is positively relevant to B). Write just „Yes‟ or „No‟ in each column provided. A B Conclusive Some evidence? evidence? (i) Fred just hit a hole-in-one Fred is a very good golfer No Yes (ii) I have between 4 and 6 eggs I have at least 3 eggs Yes (iii) I have at least 4 eggs I have 5 or more eggs No Yes (iv) It‟s not the case that every Every politician is honest No Yes politician is corrupt. (v) Janet likes listening to Jazz Vancouver is west of Calgary No No (vi) Simpson is a world-class skier Simpson is a non-smoker No Yes (vii) We‟re having fish for supper We‟re having trout for supper No Yes (viii) Qin‟s theory is rejected by all Qin‟s theory is false No Yes relevant scientific authorities (ix) Sally is a Catholic Sally has eight children No Yes (x) Ali stabbed a man in Vernon, Ali first entered Canada in No No B.C. in 1998. 2002. [1 mark each] In rows where you say ‘yes’ in the first column, you should of course say ‘yes’ in the second as well! Or just leave the second column blank, if you prefer. 2. Each of the following definitions is flawed in some way (each in just one way, I think, or at least one main one). -

Arguments from Design
Arguments from Design: A Self-defeating Strategy? Victoria S. Harrison University of Glasgow This is an archived version of ‘Arguments form Design: A Self-defeating Strategy?’, published in Philosophia 33 (2005): 297–317. Dr V. Harrison Department of Philosophy 69 Oakfield Avenue University of Glasgow Glasgow G12 8LT Scotland UK E-mail: [email protected] Arguments from Design: A Self-defeating Strategy? Abstract: In this article, after reviewing traditional arguments from design, I consider some more recent versions: the so-called ‘new design arguments’ for the existence of God. These arguments enjoy an apparent advantage over the traditional arguments from design by avoiding some of Hume’s famous criticisms. However, in seeking to render religion and science compatible, it seems that they require a modification not only of our scientific understanding but also of the traditional conception of God. Moreover, there is a key problem with arguments from design that Mill raised to which the new arguments seem no less vulnerable than the older versions. The view that science and religion are complementary has at least one significant advantage over other positions, such as the view that they are in an antagonistic relationship or the view that they are so incommensurable that they are neither complementary nor antagonistic. The advantage is that it aspires to provide a unified worldview that is sensitive to the claims of both science and religion. And surely, such a worldview, if available, would seem to be superior to one in which, say, scientific and religious claims were held despite their obvious contradictions. -

Intelligent Design Is Not Science” Given by John G
An Outline of a lecture entitled, “Intelligent Design is not Science” given by John G. Wise in the Spring Semester of 2007: Slide 1 Why… … do humans have so much trouble with wisdom teeth? … is childbirth so dangerous and painful? Because a big, thinking brain is an advantage, and evolution is imperfect. Slide 2 Charles Darwin’s Revolutionary Idea His book changed the world. All life forms on this planet are related to each other through “Descent with modification over generations from a common ancestor”. Natural processes fully explain the biological connections between all life on the planet. Slide 3 Darwin’s Idea is Dangerous (from the book of the same title by Daniel Dennett) If we have evolution, we no longer need a Creator to create each and every species. Darwinism is dangerous because it infers that God did not directly and purposefully create us. It simply states that we evolved. Slide 4 Intelligent Design Attempts to Counter Darwin “Intelligent Design” has been proposed as way out of this dilemma. – Phillip Johnson, William Dembski, and Michael Behe (to name a few). They attempt to redefine science to encompass the supernatural as well as the natural world. They accept Darwin’s evolution, if an Intelligent Designer is (sometimes) substituted for natural selection. Design arguments are not new: − 1250 - St. Thomas Aquinas - first design argument − 1802 - Natural Theology - William Paley – perhaps the best(?) design argument Slide 5 What is Science? A specific way of understanding the natural world. Based on the idea that our senses give us accurate information about the Universe. -

Intelligent Design Creationism and the Constitution
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Washington University St. Louis: Open Scholarship Washington University Law Review Volume 83 Issue 1 2005 Is It Science Yet?: Intelligent Design Creationism and the Constitution Matthew J. Brauer Princeton University Barbara Forrest Southeastern Louisiana University Steven G. Gey Florida State University Follow this and additional works at: https://openscholarship.wustl.edu/law_lawreview Part of the Constitutional Law Commons, Education Law Commons, First Amendment Commons, Religion Law Commons, and the Science and Technology Law Commons Recommended Citation Matthew J. Brauer, Barbara Forrest, and Steven G. Gey, Is It Science Yet?: Intelligent Design Creationism and the Constitution, 83 WASH. U. L. Q. 1 (2005). Available at: https://openscholarship.wustl.edu/law_lawreview/vol83/iss1/1 This Article is brought to you for free and open access by the Law School at Washington University Open Scholarship. It has been accepted for inclusion in Washington University Law Review by an authorized administrator of Washington University Open Scholarship. For more information, please contact [email protected]. Washington University Law Quarterly VOLUME 83 NUMBER 1 2005 IS IT SCIENCE YET?: INTELLIGENT DESIGN CREATIONISM AND THE CONSTITUTION MATTHEW J. BRAUER BARBARA FORREST STEVEN G. GEY* TABLE OF CONTENTS ABSTRACT ................................................................................................... 3 INTRODUCTION.................................................................................................. -

Probability, Statistics, Evolution, and Intelligent Design
Probability, Statistics, Evolution, and Intelligent Design Peter Olofsson n the last decades, arguments against Darwinian evolution An ID Hypothesis Testing Challenge to Evolution have become increasingly sophisticated, replacing Cre- In his book The Design Inference, William Dembski introduces ationism by Intelligent Design (ID) and the book of Genesis I the “explanatory filter” as a device to rule out chance expla- by biochemistry and mathematics. As arguments claiming to nations and infer design of observed phenomena. The filter be based in probability and statistics are being used to justify also appears in his book No Free Lunch, where the description the anti-evolution stance, it may be of interest to readers of differs slightly. In essence, the filter is a variation on statistical CHANCE to investigate methods and claims of ID theorists. hypothesis testing with the main difference being that it aims at ruling out chance altogether, rather than just a specified Probability, Statistics, and Evolution null hypothesis. Once all chance explanations have been ruled The theory of evolution states in part that traits of organisms out, ‘design’ is inferred. Thus, in this context, design is merely are passed on to successive generations through genetic mate- viewed as the complement of chance. rial and that modifications in genetic material cause changes To illustrate the filter, Dembski uses the example of in appearance, ability, function, and survival of organisms. Nicholas Caputo, a New Jersey Democrat who was in charge Genetic changes that are advantageous to successful repro- of putting together the ballots in his county. Names were to be duction over time dominate and new species evolve. -

Irreducible Complexity and Darwinian Gradualism: a Reply to Michael J
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Asbury Theological Seminary Faith and Philosophy: Journal of the Society of Christian Philosophers Volume 19 Issue 1 Article 1 1-1-2002 Irreducible Complexity and Darwinian Gradualism: A Reply to Michael J. Behe Paul Draper Follow this and additional works at: https://place.asburyseminary.edu/faithandphilosophy Recommended Citation Draper, Paul (2002) "Irreducible Complexity and Darwinian Gradualism: A Reply to Michael J. Behe," Faith and Philosophy: Journal of the Society of Christian Philosophers: Vol. 19 : Iss. 1 , Article 1. Available at: https://place.asburyseminary.edu/faithandphilosophy/vol19/iss1/1 This Article is brought to you for free and open access by the Journals at ePLACE: preserving, learning, and creative exchange. It has been accepted for inclusion in Faith and Philosophy: Journal of the Society of Christian Philosophers by an authorized editor of ePLACE: preserving, learning, and creative exchange. IRREDUCIBLE COMPLEXITY AND DARWINIAN GRADUALISM: A REPLY TO MICHAEL J. BEHE Paul Draper In Darwin's Black Box, Michael). Behe argues that, because certain biochemical systems are both irreducibly complex and very complex, it is extremely unlikely that they evolved gradually by Darwinian mechanisms, and so extremely likely that they were intelligently designed. I begin this paper by explaining Behe's argument and defending it against the very common but clearly mistaken charge that it is just a rehash of William Paley'S design argu ment. Then I critically discuss a number of more serious objections to the argument. I conclude that, while Behe successfully rules out some Darwinian paths to the biochemical systems he discusses, others remain open. -

Evidence for Design in Physics and Biology: from the Origin of the Universe to the Origin of Life
52 stephen c. meyer Pages 53–111 of Science and Evidence for Design in the Universe. The Proceedings of the Wethersfield Institute. Michael Behe, STEPHEN C. MEYER William A. Dembski, and Stephen C. Meyer (San Francisco: Ignatius Press, 2001. 2000 Homeland Foundation.) EVIDENCE FOR DESIGN IN PHYSICS AND BIOLOGY: FROM THE ORIGIN OF THE UNIVERSE TO THE ORIGIN OF LIFE 1. Introduction In the preceding essay, mathematician and probability theo- rist William Dembski notes that human beings often detect the prior activity of rational agents in the effects they leave behind.¹ Archaeologists assume, for example, that rational agents pro- duced the inscriptions on the Rosetta Stone; insurance fraud investigators detect certain ‘‘cheating patterns’’ that suggest intentional manipulation of circumstances rather than ‘‘natu- ral’’ disasters; and cryptographers distinguish between random signals and those that carry encoded messages. More importantly, Dembski’s work establishes the criteria by which we can recognize the effects of rational agents and distinguish them from the effects of natural causes. In brief, he shows that systems or sequences that are both ‘‘highly com- plex’’ (or very improbable) and ‘‘specified’’ are always produced by intelligent agents rather than by chance and/or physical- chemical laws. Complex sequences exhibit an irregular and improbable arrangement that defies expression by a simple formula or algorithm. A specification, on the other hand, is a match or correspondence between an event or object and an independently given pattern or set of functional requirements. As an illustration of the concepts of complexity and speci- fication, consider the following three sets of symbols: 53 54 stephen c. -

Does Intelligent Design Postulate a “Supernatural Creator?” Overview: No
Truth Sheet # 09-05 Does intelligent design postulate a “supernatural creator?” Overview: No. The ACLU, and many of its expert witnesses, have alleged that teaching the scientific theory of intelligent design (ID) is unconstitutional in all circumstances because it posits a “supernatural creator.” Yet actual statements from intelligent design theorists have made it clear that the scientific theory of intelligent design does not address metaphysical and religious questions such as the nature or identity of the designer. Firstly, the textbook being used in Dover, Of Pandas and People (Pandas ), makes it clear that design theory does not address religious or metaphysical questions, such as the nature or identity of the designer. Consider these two clear disclaimers from Pandas : "[T]he intelligent design explanation has unanswered questions of its own. But unanswered questions, which exist on both sides, are an essential part of healthy science; they define the areas of needed research. Questions often expose hidden errors that have impeded the progress of science. For example, the place of intelligent design in science has been troubling for more than a century. That is because on the whole, scientists from within Western culture failed to distinguish between intelligence, which can be recognized by uniform sensory experience, and the supernatural, which cannot. Today we recognize that appeals to intelligent design may be considered in science, as illustrated by current NASA search for extraterrestrial intelligence (SETI). Archaeology has pioneered the development of methods for distinguishing the effects of natural and intelligent causes. We should recognize, however, that if we go further, and conclude that the intelligence responsible for biological origins is outside the universe (supernatural) or within it, we do so without the help of science." 1 “[T]he concept of design implies absolutely nothing about beliefs normally associated with Christian fundamentalism, such as a young earth, a global flood, or even the existence of the Christian God. -

Comment the Communication Strategies of Neocreationism Between the United States and Europe
SISSA – International School for Advanced Studies Journal of Science Communication ISSN 1824 – 2049 http://jcom.sissa.it/ Comment The communication strategies of neocreationism between the United States and Europe Astrid Pizzo In their essay which appeared in 1972 in Models in Paleobiology , Stephen Jay Gould and Niles Eldredge, introducing the theory of punctuated equilibrium, stressed the fact that no scientific theory develops as a simple and logical extension of facts and of patiently recorded observations, and that the particular vision of the world that the scientist adheres to is able to influence, even unconsciously, the way in which data are collected, selected and then interpreted. Scientists, being aware of the existence of an intrinsic problem of prejudice in their scientific research activity, know that, in order to produce original and innovative ideas, it is fundamental to try to revolutionise their research image, to look at reality in a new light, to read data with alternative viewpoints. According to the American philosopher Robert Pennock 1,2, creationists ignore this aspect completely: they look to the Sacred Scriptures to find answers on the origin of the world and of life, and then try to interpret the empirical evidence so that it fits the scriptures. However, American creationism has changed radically in recent decades. Unlike creationists in the strict sense of the word, who use what is said in the Bible explicitly, at times even literally, to attack the theory of evolution, the advocates of Intelligent Design, who opened their season in the Seventies with the publication of Scientific Creationism by Henry Morris 3, do not adopt a stance of direct opposition to evolutionism, but try to work alongside it and to make use of scientific method to find the evidence of the divine hand in nature.