Associations Between Cryptococcus Genotypes, Phenotypes, and Clinical Parameters of Human Disease: a Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diversity of the Cryptococcus Neoformans-Cryptococcus Gattii Species Complex Marjan Bovers1, Ferry Hagen1 and Teun Boekhout1,2
S4 Rev Iberoam Micol 2008; 25: S4-S12 Review Diversity of the Cryptococcus neoformans-Cryptococcus gattii species complex Marjan Bovers1, Ferry Hagen1 and Teun Boekhout1,2 1Yeast Research Group, CBS Fungal Diversity Centre, Uppsalalaan 8, 3584 CT Utrecht, The Netherlands; 2Department of Internal Medicine and Infectious Diseases, University Medical Centre Utrecht, The Netherlands Summary More than 110 years of study of the Cryptococcus neoformans and Cryptococcus gattii species complex has resulted in an enormous accumulation of fundamental, applied biological and clinical knowledge. Recent developments in our understanding of the diversity within the species complex are presented, emphasizing the intraspecific complexity, which includes species, microspecies, hybrids, serotypes and genotypes. Each of these may have different roles in disease. An overview of obsolete and current names is presented. Key words Cryptococcus neoformans, Cryptococcus gattii, Yeast, Taxonomy, Pathogen. Diversidad del complejo de especies Cryptococcus neoformans-Cryptococcus gattii Resumen Más de 110 años de estudio sobre el complejo de especies Cryptococcus neoformans y Cryptococcus gattii han dado lugar a un gran acúmulo de conocimiento básico, aplicado y clínico. En este artículo se describen los avances recientes en la comprensión de su diversidad, haciendo énfasis en la complejidad intraespecífica que presenta y que llega a englobar especies, microespecies, híbridos, serotipos y genotipos. Cada uno de estos grupos puede jugar un papel diferente en la enfermedad. Se presenta también una visión global de los nombres obsoletos y actuales de estos taxones. Palabras clave Cryptococcus neoformans, Cryptococcus gattii, Levaduras, Patógeno, Taxonomía. Introduction racteristics of the genus Saccharomyces were not present, he placed these species in the genus Cryptococcus [166]. -

Cryptococcus Gattii Outbreak
The recent Cryptococcus gattii outbreak A deadly pathway… From Trees to Lungs to Brain ryptococcus gattii disease. The route of C has been in the infection is from breathing the news lately, due to a recent airborne organism, which outbreak in the Pacific becomes lodged in the Northwest. This new, more pulmonary tissues. virulent strain is expected to by Jay Hardy, CLS, SM (NRCM) spread further throughout the Symptoms Northwest and Northern California in the coming The pulmonary disease, Jay Hardy is the founder and months. known as cpryptococcosis, president of Hardy Diagnostics. develops slowly. After He began his career in exposure it can take two to microbiology as a Medical Technologist in Santa Barbara, twelve months for symptoms California. to appear, with the usual onset In 1980, he began at six or seven months. manufacturing culture media for the local hospitals. Today, The symptoms include: Hardy Diagnostics is the third largest media manufacturer in the U.S. • Cough that lasts weeks or months To ensure rapid and reliable turn around time, Hardy • Sharp chest pain Figure 1: An India ink stain Diagnostics maintains six • Unexplained shortness of showing the capsule surrounding distribution centers, and breath produces over 3,000 products the C. gattii cells in the yeast form used in clinical and industrial at 1,000X. Photo from Haley/CDC. • Severe headache microbiology laboratories • Confusion throughout the world. The organism is a fungus that • Fever is usually found in the soil • Night sweats www.HardyDiagnostics.com and on trees. It is not spread • Unintended weight loss from human to human or animal to human. -

A Silver Bullet in a Golden Age of Functional Genomics: the Impact of Agrobacterium-Mediated Transformation of Fungi
Idnurm, A. , Bailey, A. M., Cairns, T., Elliott, C., Foster, G., Ianiri, G., & Jeon, J. (2017). A silver bullet in a golden age of functional genomics: the impact of Agrobacterium-mediated transformation of fungi. Fungal Biology and Biotechnology, 4, [4:6]. https://doi.org/10.1186/s40694- 017-0035-0 Publisher's PDF, also known as Version of record License (if available): CC BY Link to published version (if available): 10.1186/s40694-017-0035-0 Link to publication record in Explore Bristol Research PDF-document This is the final published version of the article (version of record). It first appeared online via BMC at https://fungalbiolbiotech.biomedcentral.com/articles/10.1186/s40694-017-0035-0. Please refer to any applicable terms of use of the publisher. University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available: http://www.bristol.ac.uk/red/research-policy/pure/user-guides/ebr-terms/ Idnurm et al. Fungal Biol Biotechnol (2017) 4:6 DOI 10.1186/s40694-017-0035-0 Fungal Biology and Biotechnology REVIEW Open Access A silver bullet in a golden age of functional genomics: the impact of Agrobacterium‑mediated transformation of fungi Alexander Idnurm1* , Andy M. Bailey2, Timothy C. Cairns3, Candace E. Elliott1, Gary D. Foster2, Giuseppe Ianiri4 and Junhyun Jeon5 Abstract The implementation of Agrobacterium tumefaciens as a transformation tool revolutionized approaches to discover and understand gene functions in a large number of fungal species. A. -

Health and Disease Status in a Threatened Marsupial, the Quokka (Setonix Brachyurus)
Health and disease status in a threatened marsupial, the quokka (Setonix brachyurus) Pedro Martínez-Pérez Veterinarian (Hons), MVS Conservation Medicine This thesis was submitted in fulfilment of the requirements for the degree of Doctor of Philosophy in Wildlife Veterinary Studies, Murdoch University School of Veterinary and Life Sciences Murdoch University Murdoch, Western Australia January 2016 Declaration I declare that this thesis is my own account of my research and contains as its main content, work which has not been previously submitted for a degree at any tertiary educational institution. _______________________________________________ Pedro A. Martínez-Pérez I Abstract Between 1901 and 1931, there were at least six anecdotal records of disease outbreaks in mainland quokkas (Setonix brachyurus) that were associated with mass. This time period pre-dates the arrival of the red fox (Vulpes vulpes). Despite these outbreaks, little or no research has been carried out to establish health and disease baseline data of the fragmented and scattered, extant populations. Epidemiological data was determined for a range of potential pathogens, and established physiological reference intervals of apparently healthy, wild quokkas on Rottnest Island and mainland locations. There were significant differences between Rottnest Island and mainland quokkas. Rottnest Island animals had haemograms with mark evidence of oxidative injury and bone marrow response consistent with a regenerative normocytic hypochromic anaemia. Except alkaline phosphatase (ALP), all blood chemistry analytes where higher in mainland animals, with particular emphasis on creatine kinase (CK), alanine amino transferase (ALT), aspartate amino transferase (AST) and vitamin E. Some other key findings include a widespread presence of a novel herpesvirus (MaHV-6), the recovery of Cryptococcus neoformans var. -

A Silver Bullet in a Golden Age of Functional Genomics: the Impact of Agrobacterium-Mediated Transformation of Fungi
Idnurm et al. Fungal Biol Biotechnol (2017) 4:6 DOI 10.1186/s40694-017-0035-0 Fungal Biology and Biotechnology REVIEW Open Access A silver bullet in a golden age of functional genomics: the impact of Agrobacterium‑mediated transformation of fungi Alexander Idnurm1* , Andy M. Bailey2, Timothy C. Cairns3, Candace E. Elliott1, Gary D. Foster2, Giuseppe Ianiri4 and Junhyun Jeon5 Abstract The implementation of Agrobacterium tumefaciens as a transformation tool revolutionized approaches to discover and understand gene functions in a large number of fungal species. A. tumefaciens mediated transformation (AtMT) is one of the most transformative technologies for research on fungi developed in the last 20 years, a development arguably only surpassed by the impact of genomics. AtMT has been widely applied in forward genetics, whereby generation of strain libraries using random T-DNA insertional mutagenesis, combined with phenotypic screening, has enabled the genetic basis of many processes to be elucidated. Alternatively, AtMT has been fundamental for reverse genet- ics, where mutant isolates are generated with targeted gene deletions or disruptions, enabling gene functional roles to be determined. When combined with concomitant advances in genomics, both forward and reverse approaches using AtMT have enabled complex fungal phenotypes to be dissected at the molecular and genetic level. Addition- ally, in several cases AtMT has paved the way for the development of new species to act as models for specifc areas of fungal biology, particularly in plant pathogenic ascomycetes and in a number of basidiomycete species. Despite its impact, the implementation of AtMT has been uneven in the fungi. This review provides insight into the dynamics of expansion of new research tools into a large research community and across multiple organisms. -

Maternal–Fetal Transmission of Cryptococcus Gattii in Harbor Porpoise
LETTERS Oral Oncul, Yunus Atalay, Maternal–Fetal www.cdc.gov/EID/content/17/2/302- Yalcin Onem, Vedat Turhan, Ali appF.htm). The fi rst stomach chamber Acar, Yavuz Uyar, Transmission of contained two 3.5 cm × 2.5 cm raised, Dilek Y. Caglayik, Sezai Ozkan, Cryptococcus gattii centrally umbilicated ulcers and sev- and Levent Gorenek in Harbor Porpoise eral embedded anisakid nematodes. Author affi liations: Gulhane Military Medi- The uterus was gravid in the right cal Academy Haydarpasa Training Hospi- To the Editor: We report mater- horn with a mid-term fetus. No other tal, Istanbul, Turkey (O. Oncul, Y. Atalay, nal–fetal transmission of Cryptococ- gross lesions were identifi ed. Micro- Y. Onem, V. Turhan, A. Acar, S. Ozkan, L. cus gattii and death in a wild porpoise. scopically, the lung lesions correlated Gorenek); and Refi k Saydam National Pub- Cryptococcus neoformans and C. gat- with granulomatous to pyogranuloma- lic Health Agency, Ankara, Turkey (Y. Uyar, tii are 2 environmental, encapsulated tous infi ltrates, often with a myriad of D.Y. Caglayik) yeasts that cause invasive, potentially yeasts. life-threatening infections in humans The male fetus (length 30 cm, DOI: 10.3201/eid1702.100663 and animals (1). C. neoformans causes weight 2.4 kg), was examined sepa- disease in immunocompromised hosts, rately at a different facility than the References and C. gattii is also pathogenic in im- dam. It appeared grossly normal ex- 1. Tang YW, Li YL, Ye KL, Xu ZY, Ruo munocompetent hosts (2). Since 1999, ternally and was at a gestation of ≈5–6 SL, Fisher-Hoch SP, et al. -

12 Tremellomycetes and Related Groups
12 Tremellomycetes and Related Groups 1 1 2 1 MICHAEL WEIß ,ROBERT BAUER ,JOSE´ PAULO SAMPAIO ,FRANZ OBERWINKLER CONTENTS I. Introduction I. Introduction ................................ 00 A. Historical Concepts. ................. 00 Tremellomycetes is a fungal group full of con- B. Modern View . ........................... 00 II. Morphology and Anatomy ................. 00 trasts. It includes jelly fungi with conspicuous A. Basidiocarps . ........................... 00 macroscopic basidiomes, such as some species B. Micromorphology . ................. 00 of Tremella, as well as macroscopically invisible C. Ultrastructure. ........................... 00 inhabitants of other fungal fruiting bodies and III. Life Cycles................................... 00 a plethora of species known so far only as A. Dimorphism . ........................... 00 B. Deviance from Dimorphism . ....... 00 asexual yeasts. Tremellomycetes may be benefi- IV. Ecology ...................................... 00 cial to humans, as exemplified by the produc- A. Mycoparasitism. ................. 00 tion of edible Tremella fruiting bodies whose B. Tremellomycetous Yeasts . ....... 00 production increased in China alone from 100 C. Animal and Human Pathogens . ....... 00 MT in 1998 to more than 250,000 MT in 2007 V. Biotechnological Applications ............. 00 VI. Phylogenetic Relationships ................ 00 (Chang and Wasser 2012), or extremely harm- VII. Taxonomy................................... 00 ful, such as the systemic human pathogen Cryp- A. Taxonomy in Flow -
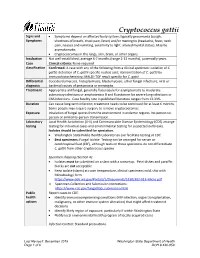
Cryptococcus Gattii Reporting and Investigation Guideline
Cryptococcus gattii Signs and • Symptoms depend on affected body system; typically pneumonia (cough, Symptoms shortness of breath, chest pain, fever) and/or meningitis (headache, fever, neck pain, nausea and vomiting, sensitivity to light, altered mental status). May be asymptomatic. • Cryptococcomas in the lungs, skin, brain, or other organs. Incubation Not well-established; average 6-7 months (range 2-13 months), potentially years. Case Clinical criteria: None required classification Confirmed: A case with any of the following from a clinical specimen: isolation of C. gattii; detection of C. gattii-specific nucleic acid; demonstration of C. gattii by immunohistochemistry; MALDI-TOF result specific for C. gattii Differential Coccidioidomycosis, histoplasmosis, blastomycosis, other fungal infections, viral or diagnosis bacterial causes of pneumonia or meningitis Treatment Appropriate antifungal, generally fluconazole for asymptomatic to moderate pulmonary infections or amphotericin B and flucytosine for severe lung infections or CNS infections. Case fatality rate in published literature ranges from 13-33%. Duration Can cause long-term infection; treatment needs to be continued for at least 6 months. Some people may require surgery to remove cryptococcomas. Exposure Inhalation of fungal spores from the environment in endemic regions. No person-to- person or animal-to-person transmission. Laboratory Local Health Jurisdiction (LHJ) and Communicable Disease Epidemiology (CDE) arrange testing testing for individual cases and environmental testing for suspected outbreaks. Isolates should be submitted for speciation. • Washington State Public Health Laboratories can facilitate testing at CDC • Best specimens: Fungal isolate. Testing can be arranged for serum or cerebrospinal fluid (CSF), although tests on these specimens do not differentiate C. gattii from other Cryptococcus species Specimen shipping (Section 4): • Isolates must be submitted on a slant with a screw top. -

Describing Genomic and Epigenomic Traits Underpinning Emerging Fungal Pathogens
1 Describing genomic and epigenomic traits underpinning 2 emerging fungal pathogens 3 4 Rhys A. Farrer1 and Matthew C. Fisher1 5 6 1Department of Infectious Disease Epidemiology, School of Public Health, Imperial 7 College London, London W2 1PG, UK 8 9 Contents 10 1. Introduction 11 2. Genome Variation Within and Between Populations of Emerging Fungal 12 Pathogens 13 2.1 Assemblies, Alignments and Annotation 14 2.2 Functional Predictions and Gene Family Expansion 15 2.3 Chromosomal Copy Number Variation 16 2.4 Natural Selection 17 2.5 Genomic approaches to detecting reproductive modes, demographic and 18 epidemiological processes in EFPs 19 2.5.1 ‘Know your enemy’ 20 2.5.2 Occurrence of EFPs caused by clonal through to reticulate 21 evolution 22 2.5.3 Mutation rates, molecular clocks and EFPs 23 3. Epigenomic Variation Within and Between Populations of Emerging Fungal 24 Pathogens 25 Acknowledgments 26 References 27 28 1 29 Abstract 30 31 An unprecedented number of pathogenic fungi are emerging and causing disease in 32 animals and plants, putting the resilience of wild and managed ecosystems in 33 jeopardy. While the past decades have seen an increase in the number of 34 pathogenic fungi, they have also seen the birth of new big-data technologies and 35 analytical approaches to tackle these emerging pathogens. We review how the 36 linked fields of genomics and epigenomics are transforming our ability to address the 37 challenge of emerging fungal pathogens. We explore the methodologies and 38 bioinformatic toolkits that currently exist to rapidly analyse the genomes of unknown 39 fungi, then discuss how these data can be used to address key questions that shed 40 light on their epidemiology. -

Cats with Central Nervous System Cryptococcosis
Acta Scientiae Veterinariae, 2020. 48(Suppl 1): 537. CASE REPORT ISSN 1679-9216 Pub. 537 Cats with Central Nervous System Cryptococcosis Rochana Rodrigues1, Jenifer Severo Beretta2, Andréia Spanamberg3, Mônica Slaviero4, Luiza Presser Ehlers4 & Luciana Sonne4 ABSTRACT Background: Cryptococcosis is a serious fungal infection contracted by humans and animals, and the most common sys- temic mycosis found in cats. This disease is often contracted through inhalation of fungal propagules. The Central Nervous System (CNS) may be infected through local extension (nasal and frontal sinuses) or via hematogenous route. Similarly to CNS bacterial infection, the clinical signs of neurological dysfunction may be attributed to mass effect (gelatinous mass of fungal microorganisms and fungal granuloma formation) or to a more disseminated inflammatory response to invading microorganisms. The objective of this study is to report one case of a patient with cryptococcal granulomas in the central nervous system and one case of a patient with neurological signs associated to a cryptococcosis. Cases: Case 1. A 3-year-old male mixed breed feline was admitted to a veterinary clinic, located in Porto Alegre, RS, Southern Brazil. The patient presented unsourced behavioral changes, vestibular ataxia and dysphagia caused by inability of coordination. The following tests were performed: complete blood count test, biochemical analysis, computed tomog- raphy scan (CT scan), fluid analysis, radiography and toxoplasmosis test. The following medicine were administrated for treatment: fluconazole, dexamethasone, mannitol, phenobarbital and levetiracetam. Fluid therapy was also part of the treatment. Immediately after death, the cat was submitted for necropsy, and a fungal granulomatous meningoencephalo- myelitis was diagnosed. Cryptococcus sp. was identified as the causal agent through pathological findings, fungal culture and PCR analysis. -

Cryptococcus Neoformans and Cryptococcus Gattii Species
Journal of Fungi Review Cryptococcus neoformans and Cryptococcus gattii Species Complexes in Latin America: A Map of Molecular Types, Genotypic Diversity, and Antifungal Susceptibility as Reported by the Latin American Cryptococcal Study Group Carolina Firacative 1,* , Wieland Meyer 2 and Elizabeth Castañeda 3 1 Studies in Translational Microbiology and Emerging Diseases (MICROS) Research Group, School of Medicine and Health Sciences, Universidad del Rosario, Bogota 111221, Colombia 2 Molecular Mycology Research Laboratory, Centre for Infectious Diseases and Microbiology, Research and Education Network Westmead Hospital, Faculty of Medicine and Health, Sydney Medical School-Westmead Clinical School, Marie Bashir Institute for Infectious Diseases and Biosecurity, Westmead Institute for Medical Research, The University of Sydney, Sydney 2145, Australia; [email protected] 3 Grupo de Microbiología, Instituto Nacional de Salud, Bogota 111321, Colombia; [email protected] * Correspondence: cfi[email protected]; Tel.: +57-1-297-0200 (ext. 3404) Abstract: Cryptococcosis, a potentially fatal mycosis, is caused by members of the Cryptococcus neoformans and Cryptococcus gattii species complexes. In Latin America, cryptococcal meningitis is still an impor- tant health threat with a significant clinical burden. Analysis of publicly available molecular data from 5686 clinical, environmental, and veterinary cryptococcal isolates from member countries of Citation: Firacative, C.; Meyer, W.; the Latin American Cryptococcal Study Group showed that, as worldwide, C. neoformans molecular Castañeda, E. Cryptococcus neoformans type VNI is the most common cause of cryptococcosis (76.01%) in HIV-infected people, followed by and Cryptococcus gattii Species C. gattii molecular type VGII (12.37%), affecting mostly otherwise healthy hosts. These two molecular Complexes in Latin America: A Map types also predominate in the environment (68.60% for VNI and 20.70% for VGII). -

3 Major Clades - Subphyla - of the Basidiomycota
3 Major Clades - Subphyla - of the Basidiomycota Agaricomycotina mushrooms, polypores, jelly fungi, corals, chanterelles, crusts, puffballs, stinkhorns Ustilaginomycotina smuts, Exobasidium, Malassezia Pucciniomycotina rusts, Septobasidium Ustilaginomycotina (Ustilaginomycetes) Ustilaginomycetes Urocystales Ustilaginales Exobasidiomycetes Exobasidiales Malasseziales Tilletiales Entorrhizomycetes simple septum with septal pore cap, not like the dolipore septum with parenthosome of Agaricomycotina Subphylum Ustilaginomycotina- smuts and relatives Ustilaginomycetes About 1500 species, 50 genera Parasitic on about 4000 spp of angiosperms, 75 families Economically important pathogens of cereals Corn smut Ustilago maydis Oat smut U. avenae Tilletia spp. “smuts and bunts” General life cycle of Ustilaginomycetes Alternate between saprobic, monokaryotic yeast and phytoparasitic, dikaryotic filamentous phases Ustilaginales-smuts • mating between monokaryotic spores • no specialized mating structures • unifactorial and bifactorial mating systems • monokaryons nonparasitic, saprobic • dikaryon phytoparasitic • heterothallic- mating of compatible spores • dimorphic- yeast and filamentous phases • teliospores teliospores germinate, give rise to a short germ tube of determinate growth called the promycelium. Promycelium: site of meiosis formation of sporidia Corn smut, Ustilago maydis Life cycle of Ustilago maydis Yeast stage, monokaryon persists in soil as saprobe Teliospores germinate to produce monokaryotic sporidia, equivalent to basidiospores Monokaryotic