Floral Resource Competition Between Honey Bees and Wild Bees: Is There Clear Evidence and Can We Guide Management and Conservation?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Conceptual Design Documentation
Appendix A: Conceptual Design Documentation APPENDIX A Conceptual Design Documentation June 2019 A-1 APPENDIX A: CONCEPTUAL DESIGN DOCUMENTATION The environmental analyses in the NEPA and CEQA documents for the proposed improvements at Oceano County Airport (the Airport) are based on conceptual designs prepared to provide a realistic basis for assessing their environmental consequences. 1. Widen runway from 50 to 60 feet 2. Widen Taxiways A, A-1, A-2, A-3, and A-4 from 20 to 25 feet 3. Relocate segmented circle and wind cone 4. Installation of taxiway edge lighting 5. Installation of hold position signage 6. Installation of a new electrical vault and connections 7. Installation of a pollution control facility (wash rack) CIVIL ENGINEERING CALCULATIONS The purpose of this conceptual design effort is to identify the amount of impervious surface, grading (cut and fill) and drainage implications of the projects identified above. The conceptual design calculations detailed in the following figures indicate that Projects 1 and 2, widening the runways and taxiways would increase the total amount of impervious surface on the Airport by 32,016 square feet, or 0.73 acres; a 6.6 percent increase in the Airport’s impervious surface area. Drainage patterns would remain the same as both the runway and taxiways would continue to sheet flow from their centerlines to the edge of pavement and then into open, grassed areas. The existing drainage system is able to accommodate the modest increase in stormwater runoff that would occur, particularly as soil conditions on the Airport are conducive to infiltration. Figure A-1 shows the locations of the seven projects incorporated in the Proposed Action. -

Bumble Bee Surveys in the Columbia River Gorge National Scenic Area of Oregon and Washington
Bumble Bee Surveys in the Columbia River Gorge National Scenic Area of Oregon and Washington Final report from the Xerces Society to the U.S. Forest Service and Interagency Special Status/Sensitive Species Program (ISSSSP) Agreement L13AC00102, Modification 5 Bombus vosnesenskii on Balsamorhiza sagittata. Photo by Rich Hatfield, the Xerces Society. By Rich Hatfield, Sarina Jepsen, and Scott Black, the Xerces Society for Invertebrate Conservation September 2017 1 Table of Contents Abstract ......................................................................................................................................................... 3 Introduction .................................................................................................................................................. 3 Methods ........................................................................................................................................................ 6 Site Selection ............................................................................................................................................. 6 Site Descriptions (west to east) ................................................................................................................ 7 T14ES27 (USFS) ..................................................................................................................................... 7 Cape Horn (USFS) ................................................................................................................................. -
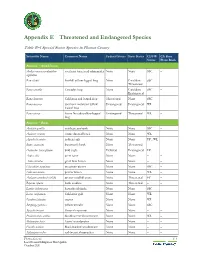
Appendix E Threatened and Endangered Species
Appendix E Threatened and Endangered Species Table E–1 Special Status Species in Plumas County Scientific Name Common Name Federal Status State Status CDFW CA Rare Status Plant Rank Animals – Amphibians Ambystoma macrodactylum southern long-toed salamander None None SSC – sigillatum Rana boylii foothill yellow-legged frog None Candidate SSC – Threatened Rana cascadae Cascades frog None Candidate SSC – Endangered Rana draytonii California red-legged frog Threatened None SSC – Rana muscosa southern mountain yellow- Endangered Endangered WL – legged frog Rana sierrae Sierra Nevada yellow-legged Endangered Threatened WL – frog Animals – Birds Accipiter gentilis northern goshawk None None SSC – Accipiter striatus sharp-shinned hawk None None WL – Aquila chrysaetos golden eagle None None FP ; WL – Buteo swainsoni Swainson's hawk None Threatened – – Haliaeetus leucocephalus bald eagle Delisted Endangered FP – Ardea alba great egret None None – – Ardea herodias great blue heron None None – – Charadrius montanus mountain plover None None SSC – Falco mexicanus prairie falcon None None WL – Antigone canadensis tabida greater sandhill crane None Threatened FP – Riparia riparia bank swallow None Threatened – – Lanius ludovicianus loggerhead shrike None None SSC – Larus californicus California gull None None WL – Pandion haliaetus osprey None None WL – Setophaga petechia yellow warbler None None SSC – Spizella breweri Brewer's sparrow None None – – Phalacrocorax auritus double-crested cormorant None None WL – Melanerpes lewis Lewis' woodpecker None -
(Hymenoptera, Apoidea, Anthophila) in Serbia
ZooKeys 1053: 43–105 (2021) A peer-reviewed open-access journal doi: 10.3897/zookeys.1053.67288 RESEARCH ARTICLE https://zookeys.pensoft.net Launched to accelerate biodiversity research Contribution to the knowledge of the bee fauna (Hymenoptera, Apoidea, Anthophila) in Serbia Sonja Mudri-Stojnić1, Andrijana Andrić2, Zlata Markov-Ristić1, Aleksandar Đukić3, Ante Vujić1 1 University of Novi Sad, Faculty of Sciences, Department of Biology and Ecology, Trg Dositeja Obradovića 2, 21000 Novi Sad, Serbia 2 University of Novi Sad, BioSense Institute, Dr Zorana Đinđića 1, 21000 Novi Sad, Serbia 3 Scientific Research Society of Biology and Ecology Students “Josif Pančić”, Trg Dositeja Obradovića 2, 21000 Novi Sad, Serbia Corresponding author: Sonja Mudri-Stojnić ([email protected]) Academic editor: Thorleif Dörfel | Received 13 April 2021 | Accepted 1 June 2021 | Published 2 August 2021 http://zoobank.org/88717A86-19ED-4E8A-8F1E-9BF0EE60959B Citation: Mudri-Stojnić S, Andrić A, Markov-Ristić Z, Đukić A, Vujić A (2021) Contribution to the knowledge of the bee fauna (Hymenoptera, Apoidea, Anthophila) in Serbia. ZooKeys 1053: 43–105. https://doi.org/10.3897/zookeys.1053.67288 Abstract The current work represents summarised data on the bee fauna in Serbia from previous publications, collections, and field data in the period from 1890 to 2020. A total of 706 species from all six of the globally widespread bee families is recorded; of the total number of recorded species, 314 have been con- firmed by determination, while 392 species are from published data. Fourteen species, collected in the last three years, are the first published records of these taxa from Serbia:Andrena barbareae (Panzer, 1805), A. -

Serves As a Model, to Show, That Solitary Bees and Their Essential Needs Must Be Considered Before the Implementation of the Measure
serves as a model, to show, that solitary bees and their essential needs must be considered before the implementation of the measure. At this habitat the characteristic heather-bees (Colletes succinctus and Andrena fuscipes) and their cuckoo bees (Epeolus cruciger and Nomada rufipes) were present. However, the local population included only a low number of individuals, with an imbalanced host-parasite relation. Maintenance measure of heather (mowing, grazing, burning or mechanical measures like “Schoppern, Abplaggen”) interfere massively with the needs of the characteristic heather-bee species and can destroy the habitats (pollen-, nectar-resources and nesting sites) for many years. The needs of the local solitary bee population must be considerate for any maintenance measure of habitats, as this case study revealed; otherwise the local population of solitary bees might diminish. Plant protection, pollination Pflanzenschutz, Bestäubung 2. BICOPOLL - Targeted precision biocontrol and pollination enhancement in organic cropping systems. Dr. Otto Boecking, Victoria Kreipe (Celle) Organic berry and fruit production suffers heavily from the lack of effective disease and pest management tools, and from inadequate insect pollination at times. As a consequence, the expanding demand on organic berries cannot be filled today. BICOPOLL expects to change this, and to significantly improve the yield and quality of organic fruit and berry production and thus, farm economics. We will use bees to (i) target deliver biological control agents to the flowers of the target crops to provide control of problem diseases, and to (ii) improve the pollination of organic horticultural crops. We will provide a pan-European case study on protecting organic strawberry from its most important disease, the grey mould. -

Eucera, Beiträge Zur Apidologie
Eucera Beiträge zur Apidologie Nr. 10 ISSN 1866-1521 25. November 2016 Eucera 10, 2016 Inhaltsverzeichnis Paul Westrich: Zum Pollensammelverhalten der auf Kreuzblütler spezialisierten Bienenarten Andrena ranunculorum und Andrena probata (Hymenoptera, Apidae) .................................................................................. 3 Paul Westrich & Josef Bülles: Epeolus fallax, ein Brutparasit von Colletes hederae und eine für Deutschland neue Bienenart (Hymenoptera, Apidae) ........................................................................................................................ 15 Paul Westrich: Die Sandbiene Andrena fulva (Hymenoptera, Apidae) als Pollensammler am Pfaffenhütchen (Euonymus europaeus) .......................................................................................................................................................... 27 Content Paul Westrich: Pollen collecting behaviour of two bee species specialized on Brassicaceae: Andrena ranun- culorum und Andrena probata (Hymenoptera, Apidae) ............................................................................................. 3 Paul Westrich & Josef Bülles: Epeolus fallax, a broodparasite of Colletes hederae and a new species for the bee fauna of Germany (Hymenoptera, Apidae) ............................................................................................ 15 Paul Westrich: Andrena fulva collects pollen from spindle (Euonymus europaeus) ......................................... 27 Anmerkung des Herausgebers Seit März -

Ukiah Western Hills Open Land Acquisition & Limited Development
Ukiah Western Hills Open Land Acquisition & Limited Development Agreement Draft Initial Study & Mitigated Negative Declaration Attachments April 16, 2021 ATTACHMENT A Existing Site Photographs Existing access road Existing water tank site Existing "house site" on one of the proposed Development Parcels ATTACHMENT B Prepared For: Michelle Irace, Planning Manager Department of Community Development 300 Seminary Avenue, Ukiah, CA 95482 APNs: 001-040-83, 157- 070-01, 157-070-02, and 003-190-01 Prepared by Jacobszoon & Associates, Inc. Alicia Ives Ringstad Senior Wildlife Biologist [email protected] Date: March 11, 2021 Updated: April 8, 2021 Biological Assessment Report Table of Contents Section 1.0: Introduction .................................................................................................................................................................. 2 Section 2.0: Regulations and Descriptions ....................................................................................................................................... 2 2.1 Regulatory Setting ................................................................................................................................................................. 2 2.2 Natural Communities and Sensitive Natural Communities .................................................................................................... 3 2.3 Special-Status Species........................................................................................................................................................... -

Assessing the Potential for Deep Learning and Computer Vision to Identify Bumble Bee Species from Images Brian J
www.nature.com/scientificreports OPEN Assessing the potential for deep learning and computer vision to identify bumble bee species from images Brian J. Spiesman1*, Claudio Gratton2, Richard G. Hatfeld3, William H. Hsu4, Sarina Jepsen3, Brian McCornack1, Krushi Patel5 & Guanghui Wang5,6 Pollinators are undergoing a global decline. Although vital to pollinator conservation and ecological research, species-level identifcation is expensive, time consuming, and requires specialized taxonomic training. However, deep learning and computer vision are providing ways to open this methodological bottleneck through automated identifcation from images. Focusing on bumble bees, we compare four convolutional neural network classifcation models to evaluate prediction speed, accuracy, and the potential of this technology for automated bee identifcation. We gathered over 89,000 images of bumble bees, representing 36 species in North America, to train the ResNet, Wide ResNet, InceptionV3, and MnasNet models. Among these models, InceptionV3 presented a good balance of accuracy (91.6%) and average speed (3.34 ms). Species-level error rates were generally smaller for species represented by more training images. However, error rates also depended on the level of morphological variability among individuals within a species and similarity to other species. Continued development of this technology for automatic species identifcation and monitoring has the potential to be transformative for the felds of ecology and conservation. To this end, we present BeeMachine, a web application that allows anyone to use our classifcation model to identify bumble bees in their own images. Bees (Hymenoptera: Anthophila) serve a critical role in most terrestrial ecosystems as pollinators of crops and natural plant communities e.g.,1–3. -

Guide to Bumble Bees of the Western United States
Bumble Bees of the Western United States By Jonathan Koch James Strange A product of the U.S. Forest Service and the Pollinator Partnership Paul Williams with funding from the National Fish and Wildlife Foundation Executive Editor Larry Stritch, Ph.D., USDA Forest Service Cover: Bombus huntii foraging. Photo Leah Lewis Executive and Managing Editor Laurie Davies Adams, The Pollinator Partnership Graphic Design and Art Direction Marguerite Meyer Administration Jennifer Tsang, The Pollinator Partnership IT Production Support Elizabeth Sellers, USGS Alphabetical Quick Reference to Species B. appositus .............110 B. frigidus ..................46 B. rufocinctus ............86 B. balteatus ................22 B. griseocollis ............90 B. sitkensis ................38 B. bifarius ..................78 B. huntii ....................66 B. suckleyi ...............134 B. californicus ..........114 B. insularis ...............126 B. sylvicola .................70 B. caliginosus .............26 B. melanopygus .........62 B. ternarius ................54 B. centralis ................34 B. mixtus ...................58 B. terricola ...............106 B. crotchii ..................82 B. morrisoni ...............94 B. vagans ...................50 B. fernaldae .............130 B. nevadensis .............18 B. vandykei ................30 B. fervidus................118 B. occidentalis .........102 B. vosnesenskii ..........74 B. flavifrons ...............42 B. pensylvanicus subsp. sonorus ....122 B. franklini .................98 2 Bumble Bees of the -

The Faunistic Drift of Apoidea in Belgium
Rasmont, Leclercq, Jacob-Remacle, Pauly & Gaspar, 1993. The faunistic drift of Apoidea in Belgium. - Page 63 Rasmont, Leclercq, Jacob-Remacle, Pauly & Gaspar, 1993. The faunistic drift of Apoidea in Belgium. - Page 64 Rasmont, P., Rasmont, Leclercq, Jacob-Remacle, Pauly & Gaspar, 1993. The faunistic drift of Apoidea in Belgium. - Page 65 J.Leclercq, A.Jacob-Remacle, A.Pauly & C.Gaspar, 1993. The faunistic drift of Apoidea in Belgium. pp. 65-87 in E. Bruneau, Bees for pollination, Commission of the European Communities, Brussels, 237 pp. THE FAUNISTIC DRIFT OF APOIDEA IN BELGIUM Pierre RASMONT*, Jean LECLERCQ**, Annie JACOB-REMACLE**, Alain PAULY** & Charles GASPAR** * Laboratoire de Zoologie Université de Mons-Hainaut Avenue Maistriau, 19 B-7000 Mons Belgique ** Zoologie générale et appliquée Faculté des Sciences agronomiques B-5030 Gembloux Belgique ABSTRACT The authors studied the faunistic drift in Apoidea of Belgium by comparing the relative number of species before and 1950 onwards. The change in the abundance was estimated by the Stroot & Depiereux (1989) method. On 360 species, 91 are decreasing (25,2%), 145 are stable (40,2%), 39 are expanding (10,8%), and 85 have an indeterminable status (rare species: 23,5%). This regression cannot be attributed to a lack of observations as they are more numerous in the second period. The authors compare different hypotheses that could explain this global regression. As the most important one affects species with a long tongue, it is likely due to the fall in availability of plants with long corollae (e.g. Lamiaceae, Fabaceae, Scrophulariaceae). The strong relative regression of cleptoparasites could be seen as the result of an absolute numerical decrease of all Apoidea. -

The Bee Fauna of the South Wales Coalfield
The Bee Fauna of the South Wales Coalfield By Liam Olds Colliery Spoil Biodiversity Initiative The South Wales coalfield is the largest continuous coalfield in Great Britain. It has an area of approximately 1000 square miles. Tylorstown Colliery, Rhondda Fach Source: Northern Mine Research Society *Please note that the data contained here is not an exhaustive list They’ve gone from this……. Tylorstown Tip, 05/08/1971 ©MaryGillhamArchives ….to this!!! Gelli Coal Tips, 16/08/2016 (Photo: Liam Olds) What is a bee? Kingdom: Animalia Phylum: Arthropoda (from Greek arthro-, joint + podos, foot) Class: Insecta (3-part body, 3 pairs of legs) Order: Hymenoptera (two pairs of membranous wings and an ovipositor specialized for stinging or piercing) Superfamily: Apoidea Family: Andrenidae, Apidae, Colletidae, Halictidae, Megachilidae, Melittidae • Around 250 species in the UK → social bees (bumblebees & the honeybee) account for around one-tenth of that figure → remainder are solitary • Quarter of bees are ‘cuckoos’ of other bee species In cuckoo solitary bees: female enters host nest and lays an egg. When the egg of the cuckoo bee hatches, the grub eats the egg/young grub of the host and uses the food provisions for its own development. Steven Falk's Flickr Collection for Hymenoptera In cuckoo bumblebees: Female enters a host nest, replaces the queen (by killing or subduing her) and then occupies the nest for the rest of its cycle. Workers become slaves. Moss carder bee (Bombus muscorum) ©Steven Falk Larger and are always covered with dense hair Vary considerably in size, appearance and where they choose to nest. Do not have pollen baskets Megachile willughbiella ©Steven Falk Andrena flavipes ©Steven Falk Some species (leafcutter and mason bees) collect pollen Most collect pollen on their legs on specialised hairs (scopa) on the underneath of their abdomen. -

(Genus: Bombus): from the Organism to the Environment AD
UNIVERSITY OF CALIFORNIA RIVERSIDE Feeding and Foraging in Bumble bees (Genus: Bombus): From the Organism to the Environment A Dissertation submitted in partial satisfaction of the requirements for the degree of Doctor of Philosophy in Entomology by Kaleigh Fisher June 2021 Dissertation Committee: Dr. S. Hollis Woodard, Chairperson Dr. Jessica Purcell Dr. Naoki Yamanaka Dr. Anupama Dahanukar Copyright by Kaleigh Fisher 2021 The Dissertation of Kaleigh Fisher is approved: Committee Chairperson University of California, Riverside Acknowledgements I would first like to thank my mentor Dr. S. Hollis Woodard for her exceptional mentorship and friendship over the course of my doctoral studies. Her support has been critical for my success as a scientist and I sincerely value all of the skills that I have developed through her mentorship. I thank her for always believing in my potential, encouraging me to pursue the exciting ideas, and showing me how to be an overall awesome scientist through her example as an amazing role model. I am also grateful for all of the members of my dissertation and qualifying exam committees Dr. Jessica Purcell, Dr. Naoki Yamanaka, Dr. Anupama Dahanukar, Dr. Quinn McFrederick, and Dr. Jason Stajich. I am especially grateful to Dr. Purcell for her encouragement and valuable conversations throughout my doctoral studies. It was such a great experience learning from her in classes as well as collaborating with her on a social insects review. I also thank her for being such an impactful role model for me. I am also especially grateful to Dr. Yamanaka and Dr. Dahanukar for collaborating on the taste projects and always offering their time, encouragement, and feedback on projects.