From the Yixian Formation in Northeastern China
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The SHRIMP U-Pb Isotope Dating of Mesozoic Volcanic from Zhangwu-Heishan Area, West Liaoning Province, China
EARTH SCIENCES RESEARCH JOURNAL GEOLOGY Earth Sci. Res. J. Vol. 24, No. 4 (December, 2020): 409-417 The SHRIMP U-Pb isotope dating of Mesozoic volcanic from Zhangwu-Heishan Area, West Liaoning Province, China Chaoyong Hou*, Houan Cai, Senlong Pei China Non-ferrous Metals Resource Geological Survey, Beijing 100012, China *Corresponding author: [email protected] ABSTRACT Keywords: West Liaoning Province; Zhangwu- The Zhangwu-Heishan area is located in the east of the Fuxin-Yixian basin. Besides the Quaternary soil, the study area Heishan Area; Mesozoic Volcanic Rock; Isotopic is mostly covered with volcanic rock. The horizon and age of volcanic rock play an essential role in understanding Age; Geological Significance. fossil beds, structures, and sedimentary evolution of West Liaoning Province and coal seeking. During this work, 11 volcanic rock samples were measured by SHRIMP U-Pb isotope analysis. Based on the reported data on the age of the Mesozoic volcanic rock in West Liaoning Province, in combination with new measurement data, Cretaceous volcanic activities in West Liaoning Province can be divided into five stages, namely 132±1 Ma, 126±1 Ma, 122±2 Ma, 115±2 Ma, and 100±5 Ma. Based on statistical results, this paper concluded that the thinning time of the crust in Northeast China is from 132±1 Ma to 115±2 Ma. Datación isotópica U-Pb a través de la Microsonda de Iones de Alta-Resolución en rocas volcánicas del Mesozoico para el área Zhangwu-Heishan, en la provincia de Liaoning, en el occidente de China RESUMEN Palabras clave: Provincia Liaoning; área Zhangwu- El área Zhangwu-Heishan se ubica en el este de la cuenca Fuxin-Yixian. -

Etude Sur L'origine Et L'évolution Des Variations Florales Chez Delphinium L. (Ranunculaceae) À Travers La Morphologie, L'anatomie Et La Tératologie
Etude sur l'origine et l'évolution des variations florales chez Delphinium L. (Ranunculaceae) à travers la morphologie, l'anatomie et la tératologie : 2019SACLS126 : NNT Thèse de doctorat de l'Université Paris-Saclay préparée à l'Université Paris-Sud ED n°567 : Sciences du végétal : du gène à l'écosystème (SDV) Spécialité de doctorat : Biologie Thèse présentée et soutenue à Paris, le 29/05/2019, par Felipe Espinosa Moreno Composition du Jury : Bernard Riera Chargé de Recherche, CNRS (MECADEV) Rapporteur Julien Bachelier Professeur, Freie Universität Berlin (DCPS) Rapporteur Catherine Damerval Directrice de Recherche, CNRS (Génétique Quantitative et Evolution Le Moulon) Présidente Dario De Franceschi Maître de Conférences, Muséum national d'Histoire naturelle (CR2P) Examinateur Sophie Nadot Professeure, Université Paris-Sud (ESE) Directrice de thèse Florian Jabbour Maître de conférences, Muséum national d'Histoire naturelle (ISYEB) Invité Etude sur l'origine et l'évolution des variations florales chez Delphinium L. (Ranunculaceae) à travers la morphologie, l'anatomie et la tératologie Remerciements Ce manuscrit présente le travail de doctorat que j'ai réalisé entre les années 2016 et 2019 au sein de l'Ecole doctorale Sciences du végétale: du gène à l'écosystème, à l'Université Paris-Saclay Paris-Sud et au Muséum national d'Histoire naturelle de Paris. Même si sa réalisation a impliqué un investissement personnel énorme, celui-ci a eu tout son sens uniquement et grâce à l'encadrement, le soutien et l'accompagnement de nombreuses personnes que je remercie de la façon la plus sincère. Je remercie très spécialement Florian Jabbour et Sophie Nadot, mes directeurs de thèse. -

Table of Codes for Each Court of Each Level
Table of Codes for Each Court of Each Level Corresponding Type Chinese Court Region Court Name Administrative Name Code Code Area Supreme People’s Court 最高人民法院 最高法 Higher People's Court of 北京市高级人民 Beijing 京 110000 1 Beijing Municipality 法院 Municipality No. 1 Intermediate People's 北京市第一中级 京 01 2 Court of Beijing Municipality 人民法院 Shijingshan Shijingshan District People’s 北京市石景山区 京 0107 110107 District of Beijing 1 Court of Beijing Municipality 人民法院 Municipality Haidian District of Haidian District People’s 北京市海淀区人 京 0108 110108 Beijing 1 Court of Beijing Municipality 民法院 Municipality Mentougou Mentougou District People’s 北京市门头沟区 京 0109 110109 District of Beijing 1 Court of Beijing Municipality 人民法院 Municipality Changping Changping District People’s 北京市昌平区人 京 0114 110114 District of Beijing 1 Court of Beijing Municipality 民法院 Municipality Yanqing County People’s 延庆县人民法院 京 0229 110229 Yanqing County 1 Court No. 2 Intermediate People's 北京市第二中级 京 02 2 Court of Beijing Municipality 人民法院 Dongcheng Dongcheng District People’s 北京市东城区人 京 0101 110101 District of Beijing 1 Court of Beijing Municipality 民法院 Municipality Xicheng District Xicheng District People’s 北京市西城区人 京 0102 110102 of Beijing 1 Court of Beijing Municipality 民法院 Municipality Fengtai District of Fengtai District People’s 北京市丰台区人 京 0106 110106 Beijing 1 Court of Beijing Municipality 民法院 Municipality 1 Fangshan District Fangshan District People’s 北京市房山区人 京 0111 110111 of Beijing 1 Court of Beijing Municipality 民法院 Municipality Daxing District of Daxing District People’s 北京市大兴区人 京 0115 -

Addition of Clopidogrel to Aspirin in 45 852 Patients with Acute Myocardial Infarction: Randomised Placebo-Controlled Trial
Articles Addition of clopidogrel to aspirin in 45 852 patients with acute myocardial infarction: randomised placebo-controlled trial COMMIT (ClOpidogrel and Metoprolol in Myocardial Infarction Trial) collaborative group* Summary Background Despite improvements in the emergency treatment of myocardial infarction (MI), early mortality and Lancet 2005; 366: 1607–21 morbidity remain high. The antiplatelet agent clopidogrel adds to the benefit of aspirin in acute coronary See Comment page 1587 syndromes without ST-segment elevation, but its effects in patients with ST-elevation MI were unclear. *Collaborators and participating hospitals listed at end of paper Methods 45 852 patients admitted to 1250 hospitals within 24 h of suspected acute MI onset were randomly Correspondence to: allocated clopidogrel 75 mg daily (n=22 961) or matching placebo (n=22 891) in addition to aspirin 162 mg daily. Dr Zhengming Chen, Clinical Trial 93% had ST-segment elevation or bundle branch block, and 7% had ST-segment depression. Treatment was to Service Unit and Epidemiological Studies Unit (CTSU), Richard Doll continue until discharge or up to 4 weeks in hospital (mean 15 days in survivors) and 93% of patients completed Building, Old Road Campus, it. The two prespecified co-primary outcomes were: (1) the composite of death, reinfarction, or stroke; and Oxford OX3 7LF, UK (2) death from any cause during the scheduled treatment period. Comparisons were by intention to treat, and [email protected] used the log-rank method. This trial is registered with ClinicalTrials.gov, number NCT00222573. or Dr Lixin Jiang, Fuwai Hospital, Findings Allocation to clopidogrel produced a highly significant 9% (95% CI 3–14) proportional reduction in death, Beijing 100037, P R China [email protected] reinfarction, or stroke (2121 [9·2%] clopidogrel vs 2310 [10·1%] placebo; p=0·002), corresponding to nine (SE 3) fewer events per 1000 patients treated for about 2 weeks. -

December 1998
JANUARY - DECEMBER 1998 SOURCE OF REPORT DATE PLACE NAME ALLEGED DS EX 2y OTHER INFORMATION CRIME Hubei Daily (?) 16/02/98 04/01/98 Xiangfan C Si Liyong (34 yrs) E 1 Sentenced to death by the Xiangfan City Hubei P Intermediate People’s Court for the embezzlement of 1,700,00 Yuan (US$20,481,9). Yunnan Police news 06/01/98 Chongqing M Zhang Weijin M 1 1 Sentenced by Chongqing No. 1 Intermediate 31/03/98 People’s Court. It was reported that Zhang Sichuan Legal News Weijin murdered his wife’s lover and one of 08/05/98 the lover’s relatives. Shenzhen Legal Daily 07/01/98 Taizhou C Zhang Yu (25 yrs, teacher) M 1 Zhang Yu was convicted of the murder of his 01/01/99 Zhejiang P girlfriend by the Taizhou City Intermediate People’s Court. It was reported that he had planned to kill both himself and his girlfriend but that the police had intervened before he could kill himself. Law Periodical 19/03/98 07/01/98 Harbin C Jing Anyi (52 yrs, retired F 1 He was reported to have defrauded some 2600 Liaoshen Evening News or 08/01/98 Heilongjiang P teacher) people out of 39 million Yuan 16/03/98 (US$4,698,795), in that he loaned money at Police Weekend News high rates of interest (20%-60% per annum). 09/07/98 Southern Daily 09/01/98 08/01/98 Puning C Shen Guangyu D, G 1 1 Convicted of the murder of three children - Guangdong P Lin Leshan (f) M 1 1 reported to have put rat poison in sugar and 8 unnamed Us 8 8 oatmeal and fed it to the three children of a man with whom she had a property dispute. -

UC Davis UC Davis Previously Published Works
UC Davis UC Davis Previously Published Works Title Integrating early Cretaceous fossils into the phylogeny of living angiosperms: Anita lines and relatives of Chloranthaceae Permalink https://escholarship.org/uc/item/3bj1s569 Journal International Journal of Plant Sciences, 175(5) ISSN 1058-5893 Authors Doyle, JA Endress, PK Publication Date 2014 DOI 10.1086/675935 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Int. J. Plant Sci. 175(5):555–600. 2014. ᭧ 2014 by The University of Chicago. All rights reserved. 1058-5893/2014/17505-0006$15.00 DOI: 10.1086/675935 INTEGRATING EARLY CRETACEOUS FOSSILS INTO THE PHYLOGENY OF LIVING ANGIOSPERMS: ANITA LINES AND RELATIVES OF CHLORANTHACEAE James A. Doyle1,* and Peter K. Endress† *Department of Evolution and Ecology, University of California, Davis, California 95616, USA; and †Institute of Systematic Botany, University of Zurich, 8008 Zurich, Switzerland Editor: Patrick S. Herendeen Premise of research. Discoveries of fossil flowers in Cretaceous rocks offer improved evidence for rela- tionships with living clades, but for more secure inferences formal phylogenetic analyses are desirable. We extend previous analyses of magnoliids, monocots, and basal eudicots to Aptian, Albian, and Cenomanian fossils related to the basal “ANITA” lines and Chloranthaceae. Methodology. We performed parsimony analyses of a morphological data set of Recent angiosperms and published fossils, with the arrangement of Recent taxa constrained to backbone trees based primarily on molecular data. Pivotal results. Not only Monetianthus (as previously inferred) but also Carpestella is nested within Nymphaeaceae, while Pluricarpellatia may be a stem relative of Cabombaceae or Nymphaeaceae. Anacostia (with Similipollis pollen) is nested within Austrobaileyales. -

Federal Register/Vol. 85, No. 12/Friday, January 17, 2020/Notices
3014 Federal Register / Vol. 85, No. 12 / Friday, January 17, 2020 / Notices public record and subject to public submissions are subject to verification, respondent selection purposes. disclosure. in accordance with section 782(i) of the Otherwise, Commerce will not collapse Tariff Act of 1930, as amended (the Act). companies for purposes of respondent Michael Fuchs, Further, in accordance with 19 CFR selection. Parties are requested to (a) Acting Director, Office of Finance and 351.303(f)(1)(i), a copy must be served identify which companies subject to Insurance Industries, Industry & Analysis, on every party on Commerce’s service review previously were collapsed, and International Trade Administration, U.S. Department of Commerce. list. (b) provide a citation to the proceeding in which they were collapsed. Further, [FR Doc. 2020–00747 Filed 1–16–20; 8:45 am] Respondent Selection if companies are requested to complete BILLING CODE 3510–DR–P In the event Commerce limits the the Quantity and Value (Q&V) number of respondents for individual Questionnaire for purposes of examination for administrative reviews DEPARTMENT OF COMMERCE respondent selection, in general, each initiated pursuant to requests made for company must report volume and value International Trade Administration the orders identified below, Commerce data separately for itself. Parties should intends to select respondents based on not include data for any other party, Initiation of Antidumping and U.S. Customs and Border Protection even if they believe they should be Countervailing Duty Administrative (CBP) data for U.S. imports during the treated as a single entity with that other Reviews POR. -
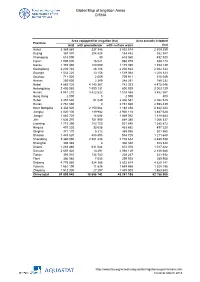
Global Map of Irrigation Areas CHINA
Global Map of Irrigation Areas CHINA Area equipped for irrigation (ha) Area actually irrigated Province total with groundwater with surface water (ha) Anhui 3 369 860 337 346 3 032 514 2 309 259 Beijing 367 870 204 428 163 442 352 387 Chongqing 618 090 30 618 060 432 520 Fujian 1 005 000 16 021 988 979 938 174 Gansu 1 355 480 180 090 1 175 390 1 153 139 Guangdong 2 230 740 28 106 2 202 634 2 042 344 Guangxi 1 532 220 13 156 1 519 064 1 208 323 Guizhou 711 920 2 009 709 911 515 049 Hainan 250 600 2 349 248 251 189 232 Hebei 4 885 720 4 143 367 742 353 4 475 046 Heilongjiang 2 400 060 1 599 131 800 929 2 003 129 Henan 4 941 210 3 422 622 1 518 588 3 862 567 Hong Kong 2 000 0 2 000 800 Hubei 2 457 630 51 049 2 406 581 2 082 525 Hunan 2 761 660 0 2 761 660 2 598 439 Inner Mongolia 3 332 520 2 150 064 1 182 456 2 842 223 Jiangsu 4 020 100 119 982 3 900 118 3 487 628 Jiangxi 1 883 720 14 688 1 869 032 1 818 684 Jilin 1 636 370 751 990 884 380 1 066 337 Liaoning 1 715 390 783 750 931 640 1 385 872 Ningxia 497 220 33 538 463 682 497 220 Qinghai 371 170 5 212 365 958 301 560 Shaanxi 1 443 620 488 895 954 725 1 211 648 Shandong 5 360 090 2 581 448 2 778 642 4 485 538 Shanghai 308 340 0 308 340 308 340 Shanxi 1 283 460 611 084 672 376 1 017 422 Sichuan 2 607 420 13 291 2 594 129 2 140 680 Tianjin 393 010 134 743 258 267 321 932 Tibet 306 980 7 055 299 925 289 908 Xinjiang 4 776 980 924 366 3 852 614 4 629 141 Yunnan 1 561 190 11 635 1 549 555 1 328 186 Zhejiang 1 512 300 27 297 1 485 003 1 463 653 China total 61 899 940 18 658 742 43 241 198 52 -

Water Lilies As Emerging Models for Darwin's Abominable Mystery
OPEN Citation: Horticulture Research (2017) 4, 17051; doi:10.1038/hortres.2017.51 www.nature.com/hortres REVIEW ARTICLE Water lilies as emerging models for Darwin’s abominable mystery Fei Chen1, Xing Liu1, Cuiwei Yu2, Yuchu Chen2, Haibao Tang1 and Liangsheng Zhang1 Water lilies are not only highly favored aquatic ornamental plants with cultural and economic importance but they also occupy a critical evolutionary space that is crucial for understanding the origin and early evolutionary trajectory of flowering plants. The birth and rapid radiation of flowering plants has interested many scientists and was considered ‘an abominable mystery’ by Charles Darwin. In searching for the angiosperm evolutionary origin and its underlying mechanisms, the genome of Amborella has shed some light on the molecular features of one of the basal angiosperm lineages; however, little is known regarding the genetics and genomics of another basal angiosperm lineage, namely, the water lily. In this study, we reviewed current molecular research and note that water lily research has entered the genomic era. We propose that the genome of the water lily is critical for studying the contentious relationship of basal angiosperms and Darwin’s ‘abominable mystery’. Four pantropical water lilies, especially the recently sequenced Nymphaea colorata, have characteristics such as small size, rapid growth rate and numerous seeds and can act as the best model for understanding the origin of angiosperms. The water lily genome is also valuable for revealing the genetics of ornamental traits and will largely accelerate the molecular breeding of water lilies. Horticulture Research (2017) 4, 17051; doi:10.1038/hortres.2017.51; Published online 4 October 2017 INTRODUCTION Ondinea, and Victoria.4,5 Floral organs differ greatly among each Ornamentals, cultural symbols and economic value family in the order Nymphaeales. -

Archaefructaceae, a New Basal Angiosperm Family
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/11380880 Archaefructaceae, a New Basal Angiosperm Family Article in Science · June 2002 DOI: 10.1126/science.1069439 · Source: PubMed CITATIONS READS 389 736 6 authors, including: Ge Sun David L Dilcher Jilin University Indiana University Bloomington 59 PUBLICATIONS 1,419 CITATIONS 393 PUBLICATIONS 13,765 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Nature and Time on Earth - Project for a course and a book for virtual visits to past environments in learning programmes for university students (coordinators Edoardo Martinetto, Emanuel Tschopp, Robert A. Gastaldo) View project NECLIME - Neogene Climate Evolution in Eurasia View project All content following this page was uploaded by David L Dilcher on 23 May 2014. The user has requested enhancement of the downloaded file. Archaefructaceae, a New Basal Angiosperm Family Ge Sun; Qiang Ji; David L. Dilcher; Shaolin Zheng; Kevin C. Nixon; Xinfu Wang Science, New Series, Vol. 296, No. 5569. (May 3, 2002), pp. 899-904. Stable URL: http://links.jstor.org/sici?sici=0036-8075%2820020503%293%3A296%3A5569%3C899%3AAANBAF%3E2.0.CO%3B2-B Science is currently published by American Association for the Advancement of Science. Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. -

First Flower 1 Have Students Bring in Flowering Plants and Distribute Them Among Groups
Original broadcast: April 17, 2007 BEFORE WATCHING First Flower 1 Have students bring in flowering plants and distribute them among groups. Ask students what the flower is for (seed formation leading PROGRAM OVERVIEW to plant reproduction). On the NOVA presents the search for the first board, draw and label the parts of a flower. Make a chart with each flowering plant. part’s function. Have students locate the following parts on their The program: flowers: anther and filament • recounts how one scientist discovered (stamen); the stigma, style, and what might be the first flowering plant ovary (pistil); and the petals fossil, Archaefructus liaoningensis. (corolla). Discuss differences in • follows the search for evidence of the first flowering plants in the reproduction between flowering and non-flowering plants. Hengdaun Mountains of China, the most biodiverse temperate forest in the world. 2 Organize students into three • states that mosses, pines, and firs dominated the Earth for 300 groups. As students watch the program, have each group take million years until flowering plants became prevalent—but how and notes on one of the following: the when this change occurred is a mystery. geography and ecology of the area • shows how flowering plants flower and produce the new featured in China; the technologies, generation—fruit—that allows the plant to adapt to a different set materials, and procedures used to study and date plant findings; and of circumstances. plant classification. • shows how Archaefructus’ separate pollen-producing organs and female organs may have evolved to be joined. • recounts the journey and discoveries of early 1900s botanist AFTER WATCHING Ernest H. -

Spermatophyta Timetree, Accelerated Base Substitution Rates at Mid
Spermatophyta timetree, accelerated base substitution rates at mid-Cretaceous and the Recent Soichi Osozawa1, Cunio Nackejima2, and john Wakabayashi3 1KawaOso Molecular Bio-Geology Institute 2Japanese Society for Plant Systematics 3California State University, Fresno September 11, 2020 Abstract We constructed a whole of Spermatophyta timetree by employing BEAST v1. X applying the nuclear ribosomal ITS, and chloroplastic matK and rbcL. Robust multipoint calibrations were done by applying fossil ages up to the Jurassic for 20 genera and a Quaternary geological event age of 1.55 Ma for 6 genera. The resultant topology was concordant to the APG system, and we successfully and precisely dated the phylogeny. Through the BEAST analyses, we discovered the exponential increase in base substitution rate in recent geologic time, and suggested that a potential cause was generation of C4 plants and the triggered Quaternary climatic change. The raised rate might have resulted in the increasing of Spermatophyta diversity including endemic Asarum and Viola species. Another rise of base substitution rate was found around 120 Ma, reflecting the order level radiation and diversification of Angiospermae at the middle Cretaceous time. Introduction A goal of botany may be the correlation of botanical evolutionary events with the timeline of Earth history (Wilf & Escapa, 2015). We prepared a Bayesian inference (BI) tree constructed using the latest and the most advanced version of BEAST (v1. X; Suchard et al ., 2018), because a credible timetree of Spermatophyta have not been constructed to date probably by employing the old version and by the unpractical use of functions (Smith et al ., 2010; Beulieu et al ., 2015).