Understanding the Dynamics in Distribution of Invasive Alien Plant Species Under Predicted Climate Change in Western Himalaya
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Food Security Bulletin 29
Nepal Food Security Bulletin Issue 29, October 2010 The focus of this edition is on the Mid and Far Western Hill and Mountain region Situation summary Figure 1. Percentage of population food insecure* 26% This Food Security Bulletin covers the period July-September and is focused on the Mid and Far Western Hill and Mountain (MFWHM) 24% region (typically the most food insecure region of the country). 22% July – August is an agricultural lean period in Nepal and typically a season of increased food insecurity. In addition, flooding and 20% landslides caused by monsoon regularly block transportation routes and result in localised crop losses. 18 % During the 2010 monsoon 1,600 families were reportedly 16 % displaced due to flooding, the Karnali Highway and other trade 14 % routes were blocked by landslides and significant crop losses were Oct -Dec Jan-M ar Apr-Jun Jul-Sep Oct -Dec Jan-M ar Apr-Jun Jul-Sep reported in Kanchanpur, Dadeldhura, western Surkhet and south- 08 09 09 09 09 10 10 10 eastern Udayapur. NeKSAP District Food Security Networks in MFWHM districts Rural Nepal Mid-Far-Western Hills&Mountains identified 163 VDCs in 12 districts that are highly food insecure. Forty-four percent of the population in Humla and Bajura are reportedly facing a high level of food insecurity. Other districts with households that are facing a high level of food insecurity are Mugu, Kalikot, Rukum, Surkhet, Achham, Doti, Bajhang, Baitadi, Dadeldhura and Darchula. These households have both very limited food stocks and limited financial resources to purchase food. Most households are coping by reducing consumption, borrowing money or food and selling assets. -

Food Security Bulletin - 21
Food Security Bulletin - 21 United Nations World Food Programme FS Bulletin, November 2008 Food Security Monitoring and Analysis System Issue 21 Highlights Over the period July to September 2008, the number of people highly and severely food insecure increased by about 50% compared to the previous quarter due to severe flooding in the East and Western Terai districts, roads obstruction because of incessant rainfall and landslides, rise in food prices and decreased production of maize and other local crops. The food security situation in the flood affected districts of Eastern and Western Terai remains precarious, requiring close monitoring, while in the majority of other districts the food security situation is likely to improve in November-December due to harvesting of the paddy crop. Decreased maize and paddy production in some districts may indicate a deteriorating food insecurity situation from January onwards. this period. However, there is an could be achieved through the provision Overview expectation of deteriorating food security of return packages consisting of food Mid and Far-Western Nepal from January onwards as in most of the and other essentials as well as A considerable improvement in food Hill and Mountain districts excessive agriculture support to restore people’s security was observed in some Hill rainfall, floods, landslides, strong wind, livelihoods. districts such as Jajarkot, Bajura, and pest diseases have badly affected In the Western Terai, a recent rapid Dailekh, Rukum, Baitadi, and Darchula. maize production and consequently assessment conducted by WFP in These districts were severely or highly reduced food stocks much below what is November, revealed that the food food insecure during April - July 2008 normally expected during this time of the security situation is still critical in because of heavy loss in winter crops, year. -

Feasibility Study of Kailash Sacred Landscape
Kailash Sacred Landscape Conservation Initiative Feasability Assessment Report - Nepal Central Department of Botany Tribhuvan University, Kirtipur, Nepal June 2010 Contributors, Advisors, Consultants Core group contributors • Chaudhary, Ram P., Professor, Central Department of Botany, Tribhuvan University; National Coordinator, KSLCI-Nepal • Shrestha, Krishna K., Head, Central Department of Botany • Jha, Pramod K., Professor, Central Department of Botany • Bhatta, Kuber P., Consultant, Kailash Sacred Landscape Project, Nepal Contributors • Acharya, M., Department of Forest, Ministry of Forests and Soil Conservation (MFSC) • Bajracharya, B., International Centre for Integrated Mountain Development (ICIMOD) • Basnet, G., Independent Consultant, Environmental Anthropologist • Basnet, T., Tribhuvan University • Belbase, N., Legal expert • Bhatta, S., Department of National Park and Wildlife Conservation • Bhusal, Y. R. Secretary, Ministry of Forest and Soil Conservation • Das, A. N., Ministry of Forest and Soil Conservation • Ghimire, S. K., Tribhuvan University • Joshi, S. P., Ministry of Forest and Soil Conservation • Khanal, S., Independent Contributor • Maharjan, R., Department of Forest • Paudel, K. C., Department of Plant Resources • Rajbhandari, K.R., Expert, Plant Biodiversity • Rimal, S., Ministry of Forest and Soil Conservation • Sah, R.N., Department of Forest • Sharma, K., Department of Hydrology • Shrestha, S. M., Department of Forest • Siwakoti, M., Tribhuvan University • Upadhyaya, M.P., National Agricultural Research Council -

Food Insecurity and Undernutrition in Nepal
SMALL AREA ESTIMATION OF FOOD INSECURITY AND UNDERNUTRITION IN NEPAL GOVERNMENT OF NEPAL National Planning Commission Secretariat Central Bureau of Statistics SMALL AREA ESTIMATION OF FOOD INSECURITY AND UNDERNUTRITION IN NEPAL GOVERNMENT OF NEPAL National Planning Commission Secretariat Central Bureau of Statistics Acknowledgements The completion of both this and the earlier feasibility report follows extensive consultation with the National Planning Commission, Central Bureau of Statistics (CBS), World Food Programme (WFP), UNICEF, World Bank, and New ERA, together with members of the Statistics and Evidence for Policy, Planning and Results (SEPPR) working group from the International Development Partners Group (IDPG) and made up of people from Asian Development Bank (ADB), Department for International Development (DFID), United Nations Development Programme (UNDP), UNICEF and United States Agency for International Development (USAID), WFP, and the World Bank. WFP, UNICEF and the World Bank commissioned this research. The statistical analysis has been undertaken by Professor Stephen Haslett, Systemetrics Research Associates and Institute of Fundamental Sciences, Massey University, New Zealand and Associate Prof Geoffrey Jones, Dr. Maris Isidro and Alison Sefton of the Institute of Fundamental Sciences - Statistics, Massey University, New Zealand. We gratefully acknowledge the considerable assistance provided at all stages by the Central Bureau of Statistics. Special thanks to Bikash Bista, Rudra Suwal, Dilli Raj Joshi, Devendra Karanjit, Bed Dhakal, Lok Khatri and Pushpa Raj Paudel. See Appendix E for the full list of people consulted. First published: December 2014 Design and processed by: Print Communication, 4241355 ISBN: 978-9937-3000-976 Suggested citation: Haslett, S., Jones, G., Isidro, M., and Sefton, A. (2014) Small Area Estimation of Food Insecurity and Undernutrition in Nepal, Central Bureau of Statistics, National Planning Commissions Secretariat, World Food Programme, UNICEF and World Bank, Kathmandu, Nepal, December 2014. -
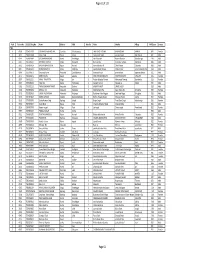
TSLC PMT Result
Page 62 of 132 Rank Token No SLC/SEE Reg No Name District Palika WardNo Father Mother Village PMTScore Gender TSLC 1 42060 7574O15075 SOBHA BOHARA BOHARA Darchula Rithachaupata 3 HARI SINGH BOHARA BIMA BOHARA AMKUR 890.1 Female 2 39231 7569013048 Sanju Singh Bajura Gotree 9 Gyanendra Singh Jansara Singh Manikanda 902.7 Male 3 40574 7559004049 LOGAJAN BHANDARI Humla ShreeNagar 1 Hari Bhandari Amani Bhandari Bhandari gau 907 Male 4 40374 6560016016 DHANRAJ TAMATA Mugu Dhainakot 8 Bali Tamata Puni kala Tamata Dalitbada 908.2 Male 5 36515 7569004014 BHUVAN BAHADUR BK Bajura Martadi 3 Karna bahadur bk Dhauli lawar Chaurata 908.5 Male 6 43877 6960005019 NANDA SINGH B K Mugu Kotdanda 9 Jaya bahadur tiruwa Muga tiruwa Luee kotdanda mugu 910.4 Male 7 40945 7535076072 Saroj raut kurmi Rautahat GarudaBairiya 7 biswanath raut pramila devi pipariya dostiya 911.3 Male 8 42712 7569023079 NISHA BUDHa Bajura Sappata 6 GAN BAHADUR BUDHA AABHARI BUDHA CHUDARI 911.4 Female 9 35970 7260012119 RAMU TAMATATA Mugu Seri 5 Padam Bahadur Tamata Manamata Tamata Bamkanda 912.6 Female 10 36673 7375025003 Akbar Od Baitadi Pancheswor 3 Ganesh ram od Kalawati od Kalauti 915.4 Male 11 40529 7335011133 PRAMOD KUMAR PANDIT Rautahat Dharhari 5 MISHRI PANDIT URMILA DEVI 915.8 Male 12 42683 7525055002 BIMALA RAI Nuwakot Madanpur 4 Man Bahadur Rai Gauri Maya Rai Ghodghad 915.9 Female 13 42758 7525055016 SABIN AALE MAGAR Nuwakot Madanpur 4 Raj Kumar Aale Magqar Devi Aale Magar Ghodghad 915.9 Male 14 42459 7217094014 SOBHA DHAKAL Dolakha GhangSukathokar 2 Bishnu Prasad Dhakal -
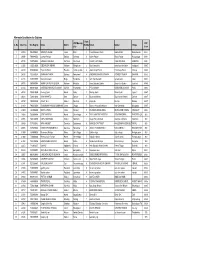
PMT Result 2075 List.Xlsx
Alternate Candidates for Diploma Ward VDC/Municip PMT S. No. Token No SLC Reg No Name District Numbe Father Mother Village ality Score r 1 29169 7063002034 NIRMAYA SHAHI Jumla Birat 3 Prem Bahadur Shahi Kabita Shahi Barkotebada 968.1 2 30698 7468080053 Laxman Pariyar Bardiya Sorhawa 9 Kuber Pariyar Bhula Pariyar Puspanagar 968.1 3 30798 7374036042 RAMESH BOHARA Darchula Sharmauli 1 HARDEV BOHARA HIRA BOHARA SARMOLI 968.1 4 33026 7100271008 SIDDHA DHUNGANA Achham Mangalsen 5 Moti Ram jaishi manshara devi jaishi kudabasti 968.2 5 33372 6724003049 Rajendra Bulun Rasuwa Laharepouwa 4 Jawan Singh Bulun Charimaya Bulun dhunge 968.2 6 28436 7429362034 SHANKAR THAPA Bajhang Patadewal 8 KRISHNA BAHADUR THAPA SITADEVI THAPA BAYANA 968.3 7 33115 7429729007 niraj kumar karki Mugu Kotdanda 6 ram chandra karki raj kala karki luwai 968.4 8 30775 7369007044 MANOJ BAHADUR BUDHA Achham Risidaha 2 Amar Bahadur Budha Dewa Devi Budha Sanikhet 968.6 9 29144 6662015026 LOKENDRA BAHADUR SINGH Dailekh Chamunda 1 PURA SINGH KARNASHILA SHAHI Palta 968.7 10 30158 7459010030 khemraj Sarki Humla Maila 2 Manrup Sarki Dhauli Sarki tajakot 968.7 11 30416 7436010092 AJAY MAHATO Bara Dahiyar 6 Bayanath Mahato Bigni Sahani Malahi Dahiyar 968.7 12 30792 7462050002 JAGAT B. K Kalikot Ranchuli 2 Anipal bk Biuri bk Ratada 968.7 13 31833 7463020058 SURENDRA PRASAD SIMKHADE Jumla Dhapa 7 Ganesh Prasad Simkhade Ram Simkhade Bistabada 968.7 14 35029 7363004002 NISHA HAMAL Jumla Narakot 2 DHARMAL BDR HAMAL DHAN LAXMI HAMAL NARAKOT 968.7 15 28683 7359004084 SITA PHADERA Humla ShreeNagar -

Notice Fifth Lot for All PMT Upto 2075.10.13 Selected & Alternate 2Nd Quintile
मत / / गतेको "गोरखाप दैनक "मा काशत सुचना अनुसार ावधक श ालयहमा डलोमा तहमा भनाx भएर वगत (चार पटक ) मा परयोजनाले सुचना काशन गदाx तो"कएको #यादभ$ आवेदन दताx गराउन छु टेका व(याथ*हले छा$वि-तृ आवेदन (Scholarship Application) फाराम परयोजनाको वेवसाइट www.event.gov.np बाट डाउनलोड गर2 भरेर श ालयले मा3णत गर2 प"रयोजना स%चवालय , वु'नगरमा मत /( /() काया* य समयभ दताx गराउनुहोला । सो5ह सुचना अनुसार डलोमा तहमा बाँक8 रहेका 99 कोटामा छा$वि-तमाृ छनोट गन: योजनका लाग पुवx काशन भएको पएमट2 यो;यता<म अनुसारको =>?@ जनाको नामावल2 । नोट : आवेदन फाराम प"रयोजनाका पुवx सुचनाह1 अनुसार दताx ग"रसकेका 2व3याथ5ह1ले भनु x नपन6 । Selected candidates for Diploma Ward Token VDC/Municipa PMT SN. SLC Reg No Name District Numb Father Mother Village No lity Score er 1 34871 7416018187 SRIJANA KUMARI MAHATO Siraha RamnagarMirchaiya 7 BINDESHWAR MAHATO SUDI RAM DULARI DEVI BAN KARYALAYA CHOWK884.5 2 28644 7259004015 DANSINGH ROKAYA Humla ShreeNagar 5 Barkhe Rokaya Gorikala Parki Rokaya Village 899 3 28719 7059004006 BANDANA PHADERA Humla ShreeNagar 4 Netra Phadera Pushi Phadera Phadera Gaun 905 4 32936 7224003027 JEEWAN KUMAR NEUPANE Rasuwa Dhaibung 4 Khem Raj Neupane Chet Kumari Neupane Katunje 907.5 5 32696 7063022017 KAMAL ROKAYA Jumla Lihi(Rara) 6 MAN BAHADUR ROKAYA MANMA ROKAYA LIHI 910 6 32817 7124013003 Ghyu Jyalmo Tamang Rasuwa Gatlang 8 Kawa Tamang Dawa Chamo Tamang Gre 910.5 7 29380 6562005014 Dipak Kumar Shahi Kalikot Jubika 6 Harsa Bahadur Shahi Padma Shahi Jubitha 911.7 8 30311 7372001067 DEEPA PARIYAR Doti Dipayal Silgadhi N.P.5 DILIP PARIYAR NIRMALA PARIYAR SILGADI 915.5 -

Danphe Newsletter (December 2011)
BirdBird ConservationConser Nepal QUARTERLY NEWSLETTER VOLUME 20 BULLETIN NO. 4 DECEMBER 2011 Impacts of agriculture on Nepal birds Carol Inskipp and Hem Sagar Baral Introduction (Himalayan Nature 2006). With support from the Oriental Bird According to BirdLife International (2008a) agriculture currently Club UK, Himalayan Nature has expanded its work on farmlands destroys and degrades more wildlife habitat than any other with permanent transects established in Koshi and Lumbini. factor worldwide resulting in biodiversity declines across It is planned to monitor these transects in early and late 2012 huge areas. Both the spread of agriculture and agricultural (Sharad Kumar Singh verbally to H. S. Baral, December 2011). intensification, such as losses in crop diversity and increases in uses of pesticides and fertilizers are to blame. Considering the damaging impacts of agriculture on bird populations in Europe and globally, a desk study was carried out In Europe data from a number of national monitoring schemes to assess the impacts on birds in Nepal in 2010. of 124 of Europe’s common bird species were analysed over the period1980-2006. This analysis showed that 45% of species Importance of agricultural lands to Nepal surveyed had declined across 20 countries, with farmland birds doing particularly badly. Intensification of agriculture was found birds The availability of invertebrates, small mammals, amphibians, to be the major cause of declines of Europe’s farmland birds spilt grain and weed seeds in cultivation provide food for a (BirdLife International, 2008b). wide range of Nepal birds. A total of 184 species, 21% of those In Nepal there has been a considerable amount of work done recorded in the country utilize agricultural habitats for feeding on globally threatened bird species in the last 20 years. -

(Naemorhedus Goral) in API NAMPA CONSERVATION AREA, NEPAL THESIS SUBMITTED to the FOREST RESEARCH INSTITUTE (DEEMED) UNIVERSITY DEHRADUN, UTTARAKHAND
DISTRIBUTION AND HABITAT PREFERENCE OF GORAL (Naemorhedus goral) IN API NAMPA CONSERVATION AREA, NEPAL THESIS SUBMITTED TO THE FOREST RESEARCH INSTITUTE (DEEMED) UNIVERSITY DEHRADUN, UTTARAKHAND FOR THE AWARD OF THE DEGREE OF MASTER OF SCIENCE IN WOOD SCIENCE AND TECHNOLOGY BY PEMBA SHERPA UNDER THE SUPERVISION OF DR. S. SATHYAKUMAR SCIENTIST-G DEPARTMENT OF ENDANGERED SPECIES MANAGEMENT WILDLIFE INSTITUTE OF INDIA. 2016 Forest Research Institute Deemed University Indian Council of Forestry Research and Education P.O.New Forest, Dehradun-248006, Uttarakhand, India 2014-2016 I DECLARATION I hereby declare that the thesis entitled “Distribution and habitat preference of Goral (Naemorhaedus goral) in Api nampa conservation area, Nepal” is a record of bonafide work carried out by me under the guidance of Dr. S. Sathyakumar, Scientist G, Wildlife Institute of India, Dehradun and no part of this work has been submitted for any other degree or diploma to any institution or university. Date: (Pemba Sherpa) Place: Enrollment No: F/14/29 II FOREST RESEARCH INSTITUTE (DEEMED) UNIVERSITY (INDIAN COUNCIL OF FORESTRY RESEARCH & EDUCATION) P.O.: I.P.E. KUALAGARH ROAD, DEHRADUN-24819. Dr. S.P.S.Rawat (Dean Academic) P.O New Forest, Dehradun 248006, INDIA E Mail: [email protected] Ph. (O) :0135-2752682 Fax : 0135-2752682 EPABX : 0135222-4452 CERTIFICATE This is to certify that the thesis entitled “Distribution and habitat preference of Goral(Naemorhaedus goral) in Api nampa conservation area, Nepal” is an original record of confide research work carried out by Mr. Pemba Sherpa, student of M.Sc. Forestry (2014-2016) of Forest Research Institute (Deemed) University, Dehradun and submitted in partial fulfillment of the requirement for M.Sc. -

A Sociolinguistic Study of Dotyali
DigitalResources Electronic Survey Report 2019-004 A Sociolinguistic Study of Dotyali Stephanie R. Eichentopf and Katharina Tupper A Sociolinguistic Study of Dotyali Stephanie R. Eichentopf and Katharina Tupper SIL International® 2019 SIL Electronic Survey Report 2019-004, May 2019 © 2019 SIL International® All rights reserved Data and materials collected by researchers in an era before documentation of permission was standardized may be included in this publication. SIL makes diligent efforts to identify and acknowledge sources and to obtain appropriate permissions wherever possible, acting in good faith and on the best information available at the time of publication. Abstract This report presents the results of sociolinguistic research conducted among Dotyali communities in west Nepal. Because of the recent ISO designation of Dotyali as an independent language, the presence of various development organizations in the region, and the overall goals of the Linguistic Survey of Nepal (LinSuN),1 basic sociolinguistic information was sought. The primary goal of this research was to gain a better understanding of the dialects of Dotyali and the level of comprehension between those dialects. Specifically, this research sought to clarify the level of comprehension of the Dotyali spoken in Doti district among speakers in other districts in order to determine their ability to share language-based materials. Language attitudes, ethnolinguistic identity, and language vitality were also addressed in this study in order to ensure a holistic picture of the sociolinguistic situation. This research identified four potential dialects of Dotyali and established that comprehension between these dialects is high. Attitudes expressed towards the Doti variety of Dotyali by speakers from other districts were positive. -

A Sociolinguistic Study of Dotyali
A Sociolinguistic Study of Dotyali Written by: Stephanie R. Eichentopf Research conducted by: Stephanie R. Eichentopf Sara A. Boon Kimberly D. Benedict Linguistic Survey of Nepal (LinSuN) Central Department of Linguistics Tribhuvan University, Nepal and SIL International 2014 Table of Contents 1 Introduction ..................................................................................................................... 8 1.1 Geography ................................................................................................................ 9 1.2 History of the people ............................................................................................... 9 1.3 Language ................................................................................................................ 11 1.4 Other nearby languages ......................................................................................... 12 1.5 Previous research and resources ............................................................................ 14 2 Purpose and Goals .......................................................................................................... 17 3 Methodology .................................................................................................................. 19 3.1 Instruments ............................................................................................................ 19 3.2 Site selection ......................................................................................................... -

The Current Food Security Qtr
Nepal Food Security Bulletin Issue 25, July - October 2009 Situation Summary • The total number of food insecure people across Nepal is Figure 1. Percentage of population food insecure estimated to be 3.7 million, this represents approximately 16.4% of the rural population. WFP Nepal is feeding 1.6 17.0% million people which has had a significant impact on reducing 2009 winter drought this figure. 16.5% • July—August is typically a period of heightened food insecurity across Nepal. This year’s lean period was particularly severe in several areas of the country due to the 2008/09 winter 16.0% drought which led to reduced household food stocks and in the worst affected areas household food shortages. 15.5% • During the coming months, short term food security should continue to improve across most of Nepal as the current 15.0% harvest of summer crops (paddy, millet and maize) will be completed. However, the longer term outlook is that food security will decline within the next 6 months as summer crop 14.5% production at a national level is expected to be generally weak. Oct-Dec 09 Jan-Mar 09 Apr-Jun 09 Jul-Sep 09 Poor summer crop production is the result of late plantation (caused by late monsoon rains) combined with erratic and generally low rainfall during the monsoon. • Of the 476 households surveyed by WFP between July and September, summer crop losses of more than 30% have been experienced or are expected by more than 40% of households. Of critical concern is the situation in Bajura, Achham, Darchula, Jumla, Humla, Mugu, Dailekh, Rukum, and Taplejung where the main summer crops (paddy,millet and/or maize) have failed by 30-70% across multiple VDCs.