Spatial and Temporal Genetic Structure of a Riverresident Atlantic Salmon (Salmo Salar) After Millennia of Isolation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Brukerundersøkelse I Raet Nasjonalpark
1606 Brukerundersøkelse i Raet nasjonalpark Sommeren 2018 Sofie K. Selvaag Line C. Wold NINAs publikasjoner NINA Rapport Dette er NINAs ordinære rapportering til oppdragsgiver etter gjennomført forsknings-, overvåkings- eller utredningsarbeid. I tillegg vil serien favne mye av instituttets øvrige rapportering, for eksempel fra seminarer og konferanser, resultater av eget forsknings- og utredningsarbeid og litteraturstudier. NINA Rapport kan også utgis på annet språk når det er hensiktsmessig.. NINA Temahefte Som navnet angir behandler temaheftene spesielle emner. Heftene utarbeides etter behov og serien favner svært vidt; fra systematiske bestemmelsesnøkler til informasjon om viktige problemstillinger i samfunnet. NINA Temahefte gis vanligvis en populærvitenskapelig form med mer vekt på illustrasjoner enn NINA Rapport. NINA Fakta Faktaarkene har som mål å gjøre NINAs forskningsresultater raskt og enkelt tilgjengelig for et større publikum. Faktaarkene gir en kort framstilling av noen av våre viktigste forskningstema. Annen publisering I tillegg til rapporteringen i NINAs egne serier publiserer instituttets ansatte en stor del av sine vitenskapelige resultater i internasjonale journaler, populærfaglige bøker og tidsskrifter. Brukerundersøkelse i Raet nasjonalpark Sommeren 2018 Sofie K. Selvaag Line C. Wold Norsk institutt for naturforskning NINA Rapport 1606 Selvaag S.K. & Wold L.C. 2019. Brukerundersøkelse i Raet nasjo- nalpark sommeren 2018. NINA Rapport 1606. Norsk institutt for na- turforskning. Lillehammer, januar 2019 ISSN: 1504-3312 -

Fiskeribiologiske Undersøkelser I Nidelva Og Gjøv I Åmli , Aust-Agder
FISKERIBIOLOGISKE UNDERSØKELSER I NIDELVA OG GJØV I ÅMLI , AUST-AGDER SIGURD KJETIL BJØRTUFT FORORD I forbindelse med det planlagte Åmli kraftverk ble Laborato- rium for ferskvannsØkologi og innlandsfiske bedt av Aust-Agder Kraftverk om å foreta fiskeribiologiske undersøkelser i Nidelva og GjØv. Naturforvaltningskandidat Sigurd Kjetil BjØrtuft ble engasjert for å arbeide med denne undersØkelsen. Han har stått ansvarlig for gjennomfØringen av hele undersØkelsen etter opplegg fra undertegnede. Feltarbeidet er utført i periodene 6.-11.juni, 24.-27.juni, 10.-14.juli og 7.-12.august, 1978. Foruten BjØrtuft har Harry Johansen, Jan-Olav Styrvold, Jørgen Brabrand og Finn Smedstad vært med på feltarbeidet. Vannanalysene er utfØrt av Eva Brorson, Avd. Limnologi, Institutt for marinbiologi og limnologi, Universitetet i Oslo. Det rettes en takk til alle som har vært engasjert og for- Øvrig konsultert i forbindelse med undersøkelsen. Oslo, 25.oktober, 1978. Svein Jakob Saltveit. INNHOLD SAMMENDRAG ............................ 3 INNLEDNING ............................ 5 OMRÅDEBESKRIVELSE ..................... 5 METODER ... .. ......................... 7 RESULTATER ............................ 12 Kjemiske målinger .................. 12 PrØvefisket ........................ 13 Ørret .............................. 14 Abbor .............................. 25 Ernæring ........................... 29 Gyteforhold ........................ 38 Parasitter ......................... 39 Sportsfiske 41 KOMMENTARER ........................... 42 LITTERATUR ........................... -

Bear News/Make PDF & CD
International Bear News Quarterly Newsletter of the International Association for Bear Research and Management (IBA) and the IUCN/SSC Bear Specialist Group February 2002 vol. 11, no. 1 Contents Council News 2 From the President 4 USA/Russia Polar Bear Treaty Support 5 Secretary's Report 6 IBA Travel Grants Due 6 IBA Grants 6 IUCN Grants for Meetings 7 Frank Cooper Craighead, Jr. Eurasia 10 Curiosity Trapped the Cat! 10 Thailand Claws 11 Pyrenees Brown Bear Restoration 13 Genetic Analysis in Austria 13 Romania: Electric Fences & Livestock Americas 15 Peru Workshop & Grassroots Success 17 Andean Bears in Ecuador 18 Venezuela: Andean Bear Project 19 Andean Bear Website 20 Western USA 22 Southwest USA 24 Northcentral USA © Lance Craighead 25 Northeast USA In Memory: Frank Cooper Craighead, Jr. 25 Florida 26 Improved Black Bear Live Trap Publications Zoo News 36 Ursus 12 Contents 27 Polar Bear Management in Central Park 37 16th E. Black Bear Workshop Proceedings Student Affairs 37 Sloth Bears 28 Opportunity Notices 37 Ursus: COYS and Black Bears Banned! 28 Your Proposal is Your Friend Events Opportunities 38 Grizzly Bears: Small Populations Workshop 31 Bear Work Wanted 38 Eighth Western Black Bear Workshop 31 Bear Biologists Needed in Florida 38 Future IBA Conferences Education/Outreach 39 14th IBA Conf. Registration & Information 32 Call for Educational Materials IBA Bears in Culture 44 Contact Information 33 Kermode Bears: From Myth to Mutation 45 Membership Application & Survey 34 Bear Self-Medication 47 Publications Order Form IBA website: www.bearbiology.com Council News From the President Harry Reynolds For example, since 1993, the IBA findings of bear biology that make Alaska Department of Fish and Game grants program (formerly the Bevins science-based conservation an 1300 College Road grant review committee) has dis- effective tool in maintaining the Fairbanks, AK 99701, USA bursed US$108,000 on 30 projects to place of bears in our world. -

Problemkartlegging Av Eutrofierte Vannforekomster I Vannområdene Gjerstad-Vegår, Nidelva Og Tovdalselva. Kunnskapsstatus Og Pr
RAPPORT L.NR. 7405-2019 Problemkartlegging av eutrofierte vannforekomster i vannområdene Gjerstad-Vegår, Nidelva og Tovdalselva. Kunnskapsstatus og prioritering av lokaliteter for screening-undersøkelser. Tovdalselva ved Høygilt, foto Jarle Håvardstun, NIVA Norsk institutt for vannforskning RAPPORT Hovedkontor NIVA Region Sør NIVA Region Innlandet NIVA Region Vest NIVA Danmark Gaustadalléen 21 Jon Lilletuns vei 3 Sandvikaveien 59 Thormøhlensgate 53 D Njalsgade 76, 4. sal 0349 Oslo 4879 Grimstad 2312 Ottestad 5006 Bergen 2300 København S, Danmark Telefon (47) 22 18 51 00 Telefon (47) 22 18 51 00 Telefon (47) 22 18 51 00 Telefon (47) 22 18 51 00 Telefon (45) 39 17 97 33 Internett: www.niva.no Tittel Løpenummer Dato Problemkartlegging av eutrofierte vannforekomster i vannområdene 7405-2019 8.07.2019 Gjerstad-Vegår, Nidelva og Tovdalselva. Kunnskapsstatus og prioritering av lokaliteter for screening-undersøkelser. Forfatter(e) Fagområde Distribusjon Hobæk, Anders Eutrofiering Åpen Geografisk område Sider Agder 66 Oppdragsgiver(e) Oppdragsreferanse Vest-Agder fylkeskommune Tanja Øverland Utgitt av NIVA Prosjektnummer 180233 Sammendrag Rapporten sammenfatter oppdatert kunnskapsstatus for 45 vannforekomster i vannområdene Gjerstad-Vegår, Nidelva og Tovdalselva. På grunnlag av tilgjengelige data er det utarbeidet forslag til hvilke vannforekomster som prioriteres for å inngå i et screening-program (enkle undersøkelser som indikerer økologisk tilstand). Prioritering av vannforekomster for screening er basert på omfang, alder og kvalitet på tilgjengelig informasjon. Som resultat av oppdatert kunnskapsstatus er 60 stasjoner for prøvetaking i screening-undersøkelser foreslått: 13 stasjoner i vannområde Gjerstad-Vegår, 33 i vannområde Nidelva og 14 i vannområde Tovdalselva. 58 stasjoner er i bekker og elver, og to stasjoner i innsjøer. -

Heimskringla III.Pdf
SNORRI STURLUSON HEIMSKRINGLA VOLUME III The printing of this book is made possible by a gift to the University of Cambridge in memory of Dorothea Coke, Skjæret, 1951 Snorri SturluSon HE iMSKrinGlA V oluME iii MAG nÚS ÓlÁFSSon to MAGnÚS ErlinGSSon translated by AliSon FinlAY and AntHonY FAulKES ViKinG SoCiEtY For NORTHErn rESEArCH uniVErSitY CollEGE lonDon 2015 © VIKING SOCIETY 2015 ISBN: 978-0-903521-93-2 The cover illustration is of a scene from the Battle of Stamford Bridge in the Life of St Edward the Confessor in Cambridge University Library MS Ee.3.59 fol. 32v. Haraldr Sigurðarson is the central figure in a red tunic wielding a large battle-axe. Printed by Short Run Press Limited, Exeter CONTENTS INTRODUCTION ................................................................................ vii Sources ............................................................................................. xi This Translation ............................................................................. xiv BIBLIOGRAPHICAL REFERENCES ............................................ xvi HEIMSKRINGLA III ............................................................................ 1 Magnúss saga ins góða ..................................................................... 3 Haralds saga Sigurðarsonar ............................................................ 41 Óláfs saga kyrra ............................................................................ 123 Magnúss saga berfœtts .................................................................. 127 -

Utlegging Av Gytegrus for Laks I Nidelva - Undersøkelse Av Gytegroper Og Ungfisk 2003-2006
LABORATORIUM FOR FERSKVANNSØKOLOGI OG INNLANDSFISKE LFI-UNIFOB UNIVERSITETET I BERGEN Rapport nr 135 Utlegging av gytegrus for laks i Nidelva - undersøkelse av gytegroper og ungfisk 2003-2006 av Bjørn T. Barlaup, Helge Skoglund, Sven-Erik Gabrielsen, Tore Wiers, Einar Kleiven og Jarle Håvardstun Etter oppdrag fra Agder-Energi Produksjon Bergen, juni 2006 LABORATORIUM FOR FERSKVANNSØKOLOGI OG INNLANDSFISKE (LFI) UNIFOB AS UNIVERSITETET I BERGEN TELEFON: 55 582228 THORMØHLENSGATE 49 TELEFAX: 55 589674 5006 BERGEN ISSN NR: LFI-RAPPORT NR: ISSN-0801-9576 135 TITTEL: DATO: Utlegging av gytegrus i Nidelva - undersøkelse av Juni 2006 gytegroper og ungfisk i 2003 - 2006 FORFATTERE: GEOGRAFISK OMRÅDE: Bjørn T. Barlaup1, Helge Skoglund1, Sven-Erik Aust-Agder Gabrielsen1, Tore Wiers1, Einar Kleiven2 og Jarle Håvardstun2 1 LFI-UNIFOB, Universitetet i Bergen 2 NIVA, Sørlandsavdelingen OPPDRAGSGIVER: ANTALL SIDER: 39 Agder Energi Produksjon AS FORSIDEFOTO: Gytegrop med lakserogn funnet i utlagt grus (venstre bilde) og område med utlagt grus (høyre bilde) i Nidelva. Foto: B.T. Barlaup 2 Innhold Sammendrag............................................................................................................................. 4 1.0 Innledning ........................................................................................................................... 7 1.1 Bakgrunn og hensikt...................................................................................................................... 7 1.2 Beskrivelse av vassdraget............................................................................................................. -

Og Sårbarhetsvurdering I Børgefjell Nasjonalpark
543 Verdi- og sårbarhetsvurdering i Børgefjell nasjonalpark - med spesielt fokus på utvalgte lokaliteter og utfordringer knyttet til ferdsel Marianne Evju Dagmar Hagen Stefan Blumentrath Nina E. Eide NINAs publikasjoner NINA Rapport Dette er en elektronisk serie fra 2005 som erstatter de tidligere seriene NINA Fagrapport, NINA Oppdragsmelding og NINA Project Report. Normalt er dette NINAs rapportering til oppdragsgiver etter gjennomført forsknings-, overvåkings- eller utredningsarbeid. I tillegg vil serien favne mye av instituttets øvrige rapportering, for eksempel fra seminarer og konferanser, resultater av eget forsk- nings- og utredningsarbeid og litteraturstudier. NINA Rapport kan også utgis på annet språk når det er hensiktsmessig. NINA Temahefte Som navnet angir behandler temaheftene spesielle emner. Heftene utarbeides etter behov og seri- en favner svært vidt; fra systematiske bestemmelsesnøkler til informasjon om viktige problemstil- linger i samfunnet. NINA Temahefte gis vanligvis en populærvitenskapelig form med mer vekt på illustrasjoner enn NINA Rapport. NINA Fakta Faktaarkene har som mål å gjøre NINAs forskningsresultater raskt og enkelt tilgjengelig for et større publikum. De sendes til presse, ideelle organisasjoner, naturforvaltningen på ulike nivå, politikere og andre spesielt interesserte. Faktaarkene gir en kort framstilling av noen av våre viktigste forsk- ningstema. Annen publisering I tillegg til rapporteringen i NINAs egne serier publiserer instituttets ansatte en stor del av sine vi- tenskapelige resultater i internasjonale journaler, populærfaglige bøker og tidsskrifter. Norsk institutt for naturforskning Verdi- og sårbarhetsvurdering i Børgefjell nasjonalpark - med spesielt fokus på utvalgte lokaliteter og utfordringer knyttet til ferdsel Marianne Evju Dagmar Hagen Stefan Blumentrath Nina E. Eide NINA Rapport 543 Evju, M., Hagen, D., Blumentrath, S. & Eide, N. -
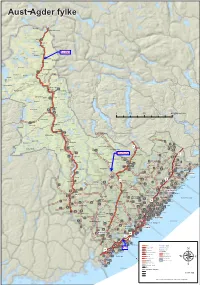
Aust−Agder Fylke
Aust−Agder fylke torhellerfjelli loros fjåen2fjellstove eggine pørsvssE fjelli freiveQRI enevssheii rovden Hartevatnet jørnrotu teinheii Vatndals− letteE vatnet tndlsE skurven dmmene Store Urevatn ferdlen Bykle ferdlsu roslemo fygdeheii Bykleheiane fyklestøyln QRP tvnes rovtn hytt Botsvatn fykle torvssE teinheii egnådlsE torsteinen heii Otra ygnestd QQT otemo uringlevtn qmsø euråhorten juven romme Raudvatnet Valleheiane tvskrdE hytt QQI lle vjomsnuten rddlsE fossu rolteheii Valle ppstd vrtenut QQR QQQ frokkeQQP ykstjørnheii rylestd 0204010 Kilometers ysstd rdvssheii wyklestøyl QQU øtefjell Øyuvsu festelnd eustd Hovatn einshornE W vngeid QPR wjåvsshytt heii qrnheim eiskvæven qjevden tkkedlen ustfjell eustd olhomfjell heii yse korv qukhei Gjøv UI ndnesÅrksø uongsfjell Bygland torrfjellet US VH ndnes qrunnetjørnsu Måvatn RI unndlsE Gjerstad ød UQ tosephsu QPQ freiung UR heii qjøvdlesklnd kåmedl hlePUP RIV QPP wosvld estøl våssen ØsterholtiIV rmreheii eustenå rdehei torlihei ylnd ndån WR QHR fyglnd Nidelva qryting romdrom wo RIU ovdl krsvssu Åmli PUI Gjavnes− UV WI undet Byglands− moen pine T vuvdl Øvre ullingsE PUU WI PUU sndre U rødneø fjorden hei PUS wjåvtn øndeled PUV ÅmliPUR egårshei Tovdalselva vuveik WQ kjeggedl festelihei RIT S QSI vongerk RIS R ivik Vegårshei F wyr IH P Barmen V WP eklnd RIT ÅrdlQPI kliknuten ippelnd fsvtn RIR Q woen isør RII W fås hølemo I qrendi RIQ xonnut øyslnd rovde ndnes Risør RIP iIV frumoen IP pie W RI ehus IHS vget fyglndsE orehei IHI tne ITR WS xipe temhei fjord vuvrk keidmo xelug PUQ IIQ RII QHP qutestdrimmelsyn -

Stabilitet Langs Namsen Utbedring Av Gamle Sikringstiltak Mot Ras Og Erosjon Anders Bjordal Mads Johnsen
Norges vassdrags- og energidirektorat Telefon: 22 95 95 95 Middelthunsgate 29 Telefaks: 22 95 90 00 Postboks 5091 Majorstua Internett: www.nve.no 0301 Oslo Stabilitet langs Namsen Utbedring av gamle sikringstiltak mot ras og erosjon Anders Bjordal Mads Johnsen 5 2007 RAPPORT Stabilitet langs Namsen Utbedring av gamle sikringstiltak mot ras og erosjon Norges vassdrags- og energidirektorat 2007 Rapport nr 5 Utbedring av gamle sikringstiltak mot ras og erosjon Utgitt av: Norges vassdrags- og energidirektorat Forfatter: Anders Bjordal, Mads Johnsen Medforfatter: Geir B Hagen, Barbro Folde, Tharan Fergus Trykk: NVEs hustrykkeri Opplag: 50 Forsidefoto: Flyfoto av Namsen ved Vibstad, anleggsarbeid på 70-tallet og anleggsarbeid i dag. Foto: NVEs fotodatabase ISBN: 978-82-410-0630-2 ISSN: 1501-2832 Sammendrag: Rapporten beskriver alle sikringsanlegg langs Namsen fra Namsos til Grong. Bakgrunnen til alle anleggene er beskrevet og alle sikringsanleggene er vist på oversiktskart. De anleggene som har behov for vedlikehold, reparasjon eller miljømessig oppgradering er beskrevet nærmere. For disse anleggene er det laget tiltaksbeskrivelse, detaljerte kart og kostnadsoverslag. Emneord: Flom, erosjon, sikringstiltak, erosjonssikringstiltak, miljøtiltak kvikkleire, kvikkleireras. Sammendrag I all tid har det gått store ras langs Namsen. Etter store skred i Namsen i 1959 og registrering av erosjonstilstanden i vassdraget i 1961, ble det i 1960 og 70-årene gjennomført et storstilt forbyggingsprogram i Namsen for å sikre mot videre erosjon og utrasing. I det store og dype elvefaret tillot ikke utstyr og arbeidsmetoder den gang en tilstrekkelige god utførelse av forbygningene. Det ble heller ikke lagt særlig vekt på istandsetting og tilrettelegging av adkomstvegen, landskap og miljø. I dag er et stort antall av de gamle forbygningene skadet og trenger reparasjon og miljømessig opprusting. -

Forekomst Av Reproduserende Bestander Av Bekke- Røye (Salvelinus Fontinalis) I Norge Pr
Forekomst av reproduserende bestander av bekke- røye (Salvelinus fontinalis) i Norge pr. 2013 Trygve Hesthagen og Einar Kleiven NINAs publikasjoner NINA Rapport Dette er en elektronisk serie fra 2005 som erstatter de tidligere seriene NINA Fagrapport, NINA Oppdragsmelding og NINA Project Report. Normalt er dette NINAs rapportering til oppdragsgiver etter gjennomført forsknings-, overvåkings- eller utredningsarbeid. I tillegg vil serien favne mye av instituttets øvrige rapportering, for eksempel fra seminarer og konferanser, resultater av eget forsk- nings- og utredningsarbeid og litteraturstudier. NINA Rapport kan også utgis på annet språk når det er hensiktsmessig. NINA Temahefte Som navnet angir behandler temaheftene spesielle emner. Heftene utarbeides etter behov og se- rien favner svært vidt; fra systematiske bestemmelsesnøkler til informasjon om viktige problemstil- linger i samfunnet. NINA Temahefte gis vanligvis en populærvitenskapelig form med mer vekt på illustrasjoner enn NINA Rapport. NINA Fakta Faktaarkene har som mål å gjøre NINAs forskningsresultater raskt og enkelt tilgjengelig for et større publikum. De sendes til presse, ideelle organisasjoner, naturforvaltningen på ulike nivå, politikere og andre spesielt interesserte. Faktaarkene gir en kort framstilling av noen av våre viktigste forsk- ningstema. Annen publisering I tillegg til rapporteringen i NINAs egne serier publiserer instituttets ansatte en stor del av sine viten- skapelige resultater i internasjonale journaler, populærfaglige bøker og tidsskrifter. Forekomst -

Project Description and Program
LINJELANGS ´along the line´ BERGEN - STAVANGER DOVREBANEN RØROSBANEN TABLE OF CONTENTS Connecting 6 villages along Sørlandsbanen.......................................... 5 HAMAR 01:00h 29 500 inhabitants Studytrip stop no. 1 BERGENSBANEN Railroad as an urbanism...................................... 7 BERGEN 08:00h 255 464 inhabitants Studytrip start The rural Experience .......................................... 9 Community railroad ............................................ 11 00:50h Nelaug................................................................ 13 OSLO 00:40h 1 000 500 inhabitants 00:45h Studytrip intersection point Linjelangs intervention........................................ 15 MOSS Participants........................................................ 17 31 000 inhabitants 00:55h Studytrip stop no. 2 PORSGRUNN 35 500 inhabitants Studytrip stop no. 3 STAVANGER 222 697 inhabitants Studytrip end 00:40h NELAUG 150 inhabitants THE SOUTHERN RAILWAY ARENDAL 44 643 inhabitants 07:45h Closed station Kristiansand 85 983 inhabitants Cities/Comunity hubs Airport 0 km 500 km CONNECTING 6 VILLAGES ALONG SØRLANDSBANEN This project aims to strengthen and revital- ize 6 rural villages along the Southern rail- road in Norway, Sørlandsbanen. To achieve this, we have constructed a two parted re- gional strategy. The first part of the strategy is strength- ening and using the existing railroad as a connection between the 6 villages. This is possible by providing a local train that op- erates between the villages. The second part is using architectural in- tervention to enhance the local resources and strengthen the connection between the village and the railroad. This strategy is based on the paradigm; that a single village is not big enough to sustain a fundamental program, but together they are big enough to function as a small scale city, with the possibility to sustain funda- mental programs. We have developed one of these villages as an example of how the regional strategy can be implemented and function in a situation. -
39428 Kystrv Namdal 05.Indd
En smak av Kystriksveien! Midtre Namdalen Namdalseid - Namsos - Fosnes - Overhalla TURISTGUIDE 2005/06 • Kart • Overnatting • Attraksjoner Innhold Kystriksveien ..................................................................... 2 Turistinformasjon/Skysstasjonen ...................................... 3 KVALITETSHYTTER OG CAMPING - HELE ÅRET Attraksjoner og aktiviteter ........................................ 5 - 34 Namsos ............................................................................... 5 Servering .................................................................... 14 NAMSOS Handel ........................................................................ 15 Overnatting ................................................................ 16 Namdalseid ...................................................................... 18 Oversikt over opplevelser i Midtre Namdalen ................ 21 Regionkart, Midtre Namdal ..................................... 22 - 23 Bykart, Namsos ................................................................ 24 Overhalla ..........................................................................25 Overnatting / Laksefiske ...........................................28 Laksefiske / Handel ...................................................30 Fosnes .............................................................................. 31 Kåret til årets Jakt og fiske ..............................................................35 - 38 camping 2004 Vandring .........................................................................