Taxonomic Revision of Trisetum Section Acrospelion
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
A REVISION of TRISETUM Victor L. Finot,' Paul M
A REVISION OF TRISETUM Victor L. Finot,' Paul M. Peterson,3 (POACEAE: POOIDEAE: Fernando 0 Zuloaga,* Robert J. v sorene, and Oscar Mattnei AVENINAE) IN SOUTH AMERICA1 ABSTRACT A taxonomic treatment of Trisetum Pers. for South America, is given. Eighteen species and six varieties of Trisetum are recognized in South America. Chile (14 species, 3 varieties) and Argentina (12 species, 5 varieties) have the greatest number of taxa in the genus. Two varieties, T. barbinode var. sclerophyllum and T longiglume var. glabratum, are endemic to Argentina, whereas T. mattheii and T nancaguense are known only from Chile. Trisetum andinum is endemic to Ecuador, T. macbridei is endemic to Peru, and T. foliosum is endemic to Venezuela. A total of four species are found in Ecuador and Peru, and there are two species in Venezuela and Colombia. The following new species are described and illustrated: Trisetum mattheii Finot and T nancaguense Finot, from Chile, and T pyramidatum Louis- Marie ex Finot, from Chile and Argentina. The following two new combinations are made: T barbinode var. sclerophyllum (Hack, ex Stuck.) Finot and T. spicatum var. cumingii (Nees ex Steud.) Finot. A key for distinguishing the species and varieties of Trisetum in South America is given. The names Koeleria cumingii Nees ex Steud., Trisetum sect. Anaulacoa Louis-Marie, Trisetum sect. Aulacoa Louis-Marie, Trisetum subg. Heterolytrum Louis-Marie, Trisetum subg. Isolytrum Louis-Marie, Trisetum subsect. Koeleriformia Louis-Marie, Trisetum subsect. Sphenopholidea Louis-Marie, Trisetum ma- lacophyllum Steud., Trisetum variabile E. Desv., and Trisetum variabile var. virescens E. Desv. are lectotypified. Key words: Aveninae, Gramineae, Poaceae, Pooideae, Trisetum. -

Investment Projects of the Republic of Dagestan Index
INVESTMENT PROJECTS OF THE REPUBLIC OF DAGESTAN INDEX INNOVATION Construction of a round and shaped steel tubes ............................. 00 producing plant Construction of the “Mountain Resources” .........................................00 Development of in-car electronics manufacturing .........................00 education and display center in Makhachkala (audio sets, starters, alternators) Construction of an IT-park of complete ............................................... 00 Construction of the “Viaduk” customs ..................................................00 “idea-series” cycle type and logistics centre Development of high-effi ciency .............................................................00 Reconstruction of the Makhachkala ..................................................... 00 solar cells and modules production commercial sea port (facilities of the second stage) Construction of the KamAZ vehicles trade ......................................... 00 INDUSTRY AND TRANSPORT and service centers in the districts of the Republic of Dagestan Development of fl oat glass production............................................... 00 Investment sites ...........................................................................................00 Development of nitric and sulfuric acid, .............................................00 and high analysis fertilizer production FUEL AND ENERGY COMPLEX onsite the “Dagfos” OJSC – II stage Construction of an intra-zone .................................................................00 -

The Vascular Plants of Massachusetts
The Vascular Plants of Massachusetts: The Vascular Plants of Massachusetts: A County Checklist • First Revision Melissa Dow Cullina, Bryan Connolly, Bruce Sorrie and Paul Somers Somers Bruce Sorrie and Paul Connolly, Bryan Cullina, Melissa Dow Revision • First A County Checklist Plants of Massachusetts: Vascular The A County Checklist First Revision Melissa Dow Cullina, Bryan Connolly, Bruce Sorrie and Paul Somers Massachusetts Natural Heritage & Endangered Species Program Massachusetts Division of Fisheries and Wildlife Natural Heritage & Endangered Species Program The Natural Heritage & Endangered Species Program (NHESP), part of the Massachusetts Division of Fisheries and Wildlife, is one of the programs forming the Natural Heritage network. NHESP is responsible for the conservation and protection of hundreds of species that are not hunted, fished, trapped, or commercially harvested in the state. The Program's highest priority is protecting the 176 species of vertebrate and invertebrate animals and 259 species of native plants that are officially listed as Endangered, Threatened or of Special Concern in Massachusetts. Endangered species conservation in Massachusetts depends on you! A major source of funding for the protection of rare and endangered species comes from voluntary donations on state income tax forms. Contributions go to the Natural Heritage & Endangered Species Fund, which provides a portion of the operating budget for the Natural Heritage & Endangered Species Program. NHESP protects rare species through biological inventory, -

Conserving Europe's Threatened Plants
Conserving Europe’s threatened plants Progress towards Target 8 of the Global Strategy for Plant Conservation Conserving Europe’s threatened plants Progress towards Target 8 of the Global Strategy for Plant Conservation By Suzanne Sharrock and Meirion Jones May 2009 Recommended citation: Sharrock, S. and Jones, M., 2009. Conserving Europe’s threatened plants: Progress towards Target 8 of the Global Strategy for Plant Conservation Botanic Gardens Conservation International, Richmond, UK ISBN 978-1-905164-30-1 Published by Botanic Gardens Conservation International Descanso House, 199 Kew Road, Richmond, Surrey, TW9 3BW, UK Design: John Morgan, [email protected] Acknowledgements The work of establishing a consolidated list of threatened Photo credits European plants was first initiated by Hugh Synge who developed the original database on which this report is based. All images are credited to BGCI with the exceptions of: We are most grateful to Hugh for providing this database to page 5, Nikos Krigas; page 8. Christophe Libert; page 10, BGCI and advising on further development of the list. The Pawel Kos; page 12 (upper), Nikos Krigas; page 14: James exacting task of inputting data from national Red Lists was Hitchmough; page 16 (lower), Jože Bavcon; page 17 (upper), carried out by Chris Cockel and without his dedicated work, the Nkos Krigas; page 20 (upper), Anca Sarbu; page 21, Nikos list would not have been completed. Thank you for your efforts Krigas; page 22 (upper) Simon Williams; page 22 (lower), RBG Chris. We are grateful to all the members of the European Kew; page 23 (upper), Jo Packet; page 23 (lower), Sandrine Botanic Gardens Consortium and other colleagues from Europe Godefroid; page 24 (upper) Jože Bavcon; page 24 (lower), Frank who provided essential advice, guidance and supplementary Scumacher; page 25 (upper) Michael Burkart; page 25, (lower) information on the species included in the database. -

Editorial 1 Liebe Leserinnen, Liebe Leser, 2 Aktuelles 6 DAV
Editorial 1 2 Aktuelles Liebe Leserinnen, liebe Leser, 2 Neue Tür für die Kreuz bachhütte 3 Jugendleiter gesucht die Abende werden früh dunkel, die mühsam 4 Ganz nah dran an den Himalaja-Riesen erworbene Sommerbräune ist schon fast völlig 5 Bremer DAV-Team auf Pionierkurs verflogen, da könnte doch die Vorstellung von einer schönen Sommerbergtour richtig wärmen. 6 DAV-unterwegs Voilà, hier vor Ihnen liegt eine wunderbare Aus- 6 Wandertouren am wahl! Unsere Tourenleiter haben auch für 2014 Monte Baldo/Gardasee 2013 wieder ein tolles Angebot zusammengestellt. 10 Naturparks im Piemont – Sie finden dieses Programm sowie Ergänzungen die Alpen am Mittelmeer und aktuelle Infos dazu jetzt auch immer im 12 Gipfel, Schluchten & Pilgerpfade Internet auf www.alpenverein-bremen.de unter 14 Berlin – 66-Seen-Wanderung der Rubrik „aktiv sein“. Im Netz können und sollten Sie auch den Newsletter abonnieren, 15 Vorträge damit sind Sie immer auf dem Laufenden. Was den treuen Lesern von BremenAlpin 16 Verein Intern spätes tens bei diesem Heft aufgefallen sein 16 Umstellung auf das dürfte: Für das Editorial schreibt jetzt jedes Mal SEPA-Lastschriftverfahren ein anderes Mitglied aus dem Vorstand. Damit 17 Günther Willenbrock verstorben ist gewährleistet, dass alle handelnden Perso- 17 Wir gratulieren zum Geburtstag nen bekannter werden und verschiedene Per- spektiven gesichert sind. 18 Für Sie gelesen Meine Sicht als 2. Vorsitzender auf die Dinge ist die, dass zurzeit viel Bewegung im 21 Tourenprogramm 2014 Verein ist. Wir sind sehr bestrebt, unsere Service- 22 Übersicht leistungen für alle Mitglieder zu verbessern. 24 Winter Hierbei hilft uns v. a. unsere Website, aber auch 25 Wandern unser Personal in der Geschäftsstelle. -

Molecular Phylogenetic Analysis Resolves Trisetum
Journal of Systematics JSE and Evolution doi: 10.1111/jse.12523 Research Article Molecular phylogenetic analysis resolves Trisetum (Poaceae: Pooideae: Koeleriinae) polyphyletic: Evidence for a new genus, Sibirotrisetum and resurrection of Acrospelion Patricia Barberá1,3*,RobertJ.Soreng2 , Paul M. Peterson2* , Konstantin Romaschenko2 , Alejandro Quintanar1, and Carlos Aedo1 1Department of Biodiversity and Conservation, Real Jardín Botánico, CSIC, Madrid 28014, Spain 2Department of Botany, National Museum of Natural History, Smithsonian Institution, Washington DC 20013‐7012, USA 3Department of Africa and Madagascar, Missouri Botanical Garden, St. Louis, MO 63110, USA *Authors for correspondence. Patricia Barberá. E‐mail: [email protected]; Paul M. Peterson. E‐mail: [email protected] Received 4 March 2019; Accepted 5 May 2019; Article first published online 22 June 2019 Abstract To investigate the evolutionary relationships among the species of Trisetum and other members of subtribe Koeleriinae, a phylogeny based on DNA sequences from four gene regions (ITS, rpl32‐trnL spacer, rps16‐trnK spacer, and rps16 intron) is presented. The analyses, including type species of all genera in Koeleriinae (Acrospelion, Avellinia, Cinnagrostis, Gaudinia, Koeleria, Leptophyllochloa, Limnodea, Peyritschia, Rostraria, Sphenopholis, Trisetaria, Trisetopsis, Trisetum), along with three outgroups, confirm previous indications of extensive polyphyly of Trisetum. We focus on the monophyletic Trisetum sect. Sibirica cladethatweinterprethereasadistinctgenus,Sibirotrisetum gen. nov. We include adescriptionofSibirotrisetum with the following seven new combinations: Sibirotrisetum aeneum, S. bifidum, S. henryi, S. scitulum, S. sibiricum, S. sibiricum subsp. litorale,andS. turcicum; and a single new combination in Acrospelion: A. distichophyllum. Trisetum s.s. is limited to one, two or three species, pending further study. Key words: Acrospelion, Aveneae, grasses, molecular systematics, Poeae, Sibirotrisetum, taxonomy, Trisetum. -

Programma 2012 in 6.0 Per MAC.Qxd 16-01-2012 12:17 Pagina 8
Programma 2012 in 6.0 per MAC.qxd 16-01-2012 12:17 Pagina 8 8 Attività 2012 Pachacamac: finalmente un tetto degno di tale nome! resoconto del Natale Alpino 2010 Il raccolto del “Natale Alpino 2010” tra i soci è andato alla comunità Raytos de luz di Pachacamac (Lima), in Perù, che ospita i bambini di strada. Ci erava- mo impegnati al rifacimento completo del tetto della loro casa con una soletta che evitasse le infiltrazioni d’acqua e permettesse in futuro l’ampliamento della struttura con un piano superiore. Grazie alla generosità dei soci ci siamo riusciti. Con la fine di marzo 2011, al termine del periodo di rinnovo tesseramento, abbiamo complessiva- I lavori sono iniziati lunedì 7 marzo 2011, mente raccolto la somma di oltre 6.600 che qui coincide con la fine dell’estate. La euro che abbiamo fatto arrivare in Perù nostra preoccupazione era infatti di riuscire a per il tramite di Alessandra Donatelli terminare i lavori prima degli inizi dell’inverno, dell’Associazione di volontariato che qui è più mite che in Italia, ma comunque “Sinergia” di Verona che ci aveva sotto- risulta piuttosto umido e i ragazzi non stareb- posto il progetto e che, come da impe- bero a loro agio negli ambienti prefabbricati di gni presi, ne ha curato e garantito la cartongesso e lamiera in cui si sono momen- realizzazione. taneamente sistemati. A fine aprile abbiamo ricevuto le foto I muratori hanno proceduto innanzitutto e la lettera che rende conto dei lavori alla demolizione della precedente copertura, eseguiti. La pubblichiamo con piacere, un multistrato leggero di canne e pali di quale atto di doveroso ringraziamento bambù, teloni di plastica e strato sottile di verso tutti i soci e gli amici (Coro Stella cemento. -
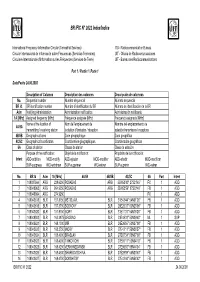
BR IFIC N° 2622 Index/Indice
BR IFIC N° 2622 Index/Indice International Frequency Information Circular (Terrestrial Services) ITU - Radiocommunication Bureau Circular Internacional de Información sobre Frecuencias (Servicios Terrenales) UIT - Oficina de Radiocomunicaciones Circulaire Internationale d'Information sur les Fréquences (Services de Terre) UIT - Bureau des Radiocommunications Part 1 / Partie 1 / Parte 1 Date/Fecha 24.06.2008 Description of Columns Description des colonnes Descripción de columnas No. Sequential number Numéro séquenciel Número sequencial BR Id. BR identification number Numéro d'identification du BR Número de identificación de la BR Adm Notifying Administration Administration notificatrice Administración notificante 1A [MHz] Assigned frequency [MHz] Fréquence assignée [MHz] Frecuencia asignada [MHz] Name of the location of Nom de l'emplacement de Nombre del emplazamiento de 4A/5A transmitting / receiving station la station d'émission / réception estación transmisora / receptora 4B/5B Geographical area Zone géographique Zona geográfica 4C/5C Geographical coordinates Coordonnées géographiques Coordenadas geográficas 6A Class of station Classe de station Clase de estación Purpose of the notification: Objet de la notification: Propósito de la notificación: Intent ADD-addition MOD-modify ADD-ajouter MOD-modifier ADD-añadir MOD-modificar SUP-suppress W/D-withdraw SUP-supprimer W/D-retirer SUP-suprimir W/D-retirar No. BR Id Adm 1A [MHz] 4A/5A 4B/5B 4C/5C 6A Part Intent 1 108037564 ARG 228.6250 POSADAS ARG 55W53'40'' 27S21'45'' FX 1 ADD 2 108048063 -

Central Asia-Caucasus
Central Asia-Caucasus Analyst BI-WEEKLY BRIEFING VOL. 12 NO. 21 10 November 2010 Searchable Archives with over 1,500 articles at http://www.cacianalyst.org ANALYTICAL ARTICLES: FIELD REPORTS: GEORGIA’S CONSTITUTIONAL REFORM Cory Welt TAJIKISTAN SEEKS TO CURB ISLAMIZATION Suhrob Majidov BP’S NEW DEAL WITH AZERBAIJAN KAZAKH PRESIDENT MAKES HISTORIC VISIT AND ITS SIGNIFICANCE TO EUROPEAN UNION Stephen Blank Georgiy Voloshin NATO’S AFGHAN SUPPLY LINES ARMENIA’S PRIME MINISTER VISITS IRAN UNDER THREAT Haroutiun Khachatrian Richard Weitz TAJIKISTAN AND UZBEKISTAN CLASH OVER NEW POLITICAL RISKS ARISE WITH THE RAILWAY TRANSIT Erkin Akhmadov NEXT GENERATIONOF METALS MINING IN KAZAKHSTAN J. Edward Conway NEWS DIGEST Central Asia-Caucasus Analyst BI-WEEKLY BRIEFING VOL. 12 NO. 21 10 NOVEMBER 2010 Contents Analytical Articles GEORGIA’S CONSTITUTIONAL REFORM 3 Cory Welt BP’S NEW DEAL WITH AZERBAIJAN AND ITS SIGNIFICANCE 6 Stephen Blank NATO’S AFGHAN SUPPLY LINES UNDER THREAT 9 Richard Weitz NEW POLITICAL RISKS ARISE WITH THE NEXT GENERATION 12 OF METALS MINING IN KAZAKHSTAN J. Edward Conway Field Reports TAJIKISTAN SEEKS TO CURB ISLAMIZATION 15 Suhrob Majidov KAZAKH PRESIDENT MAKES HISTORIC VISIT TO EUROPEAN UNION 16 Georgiy Voloshin ARMENIA’S PRIME MINISTER VISITS IRAN 17 Haroutiun Khachatrian TAJIKISTAN AND UZBEKISTAN CLASH OVER RAILWAY TRANSIT 19 Erkin Akhmadov News Digest 21 THE CENTRAL ASIA-CAUCASUS ANALYST Editor: Svante E. Cornell Associate Editor: Niklas Nilsson Assistant Editor, News Digest: Alima Bissenova Chairman, Editorial Board: S. Frederick Starr The Central Asia-Caucasus Analyst is an English-language journal devoted to analysis of the current issues facing Central Asia and the Caucasus. -

Flora Mediterranea 26
FLORA MEDITERRANEA 26 Published under the auspices of OPTIMA by the Herbarium Mediterraneum Panormitanum Palermo – 2016 FLORA MEDITERRANEA Edited on behalf of the International Foundation pro Herbario Mediterraneo by Francesco M. Raimondo, Werner Greuter & Gianniantonio Domina Editorial board G. Domina (Palermo), F. Garbari (Pisa), W. Greuter (Berlin), S. L. Jury (Reading), G. Kamari (Patras), P. Mazzola (Palermo), S. Pignatti (Roma), F. M. Raimondo (Palermo), C. Salmeri (Palermo), B. Valdés (Sevilla), G. Venturella (Palermo). Advisory Committee P. V. Arrigoni (Firenze) P. Küpfer (Neuchatel) H. M. Burdet (Genève) J. Mathez (Montpellier) A. Carapezza (Palermo) G. Moggi (Firenze) C. D. K. Cook (Zurich) E. Nardi (Firenze) R. Courtecuisse (Lille) P. L. Nimis (Trieste) V. Demoulin (Liège) D. Phitos (Patras) F. Ehrendorfer (Wien) L. Poldini (Trieste) M. Erben (Munchen) R. M. Ros Espín (Murcia) G. Giaccone (Catania) A. Strid (Copenhagen) V. H. Heywood (Reading) B. Zimmer (Berlin) Editorial Office Editorial assistance: A. M. Mannino Editorial secretariat: V. Spadaro & P. Campisi Layout & Tecnical editing: E. Di Gristina & F. La Sorte Design: V. Magro & L. C. Raimondo Redazione di "Flora Mediterranea" Herbarium Mediterraneum Panormitanum, Università di Palermo Via Lincoln, 2 I-90133 Palermo, Italy [email protected] Printed by Luxograph s.r.l., Piazza Bartolomeo da Messina, 2/E - Palermo Registration at Tribunale di Palermo, no. 27 of 12 July 1991 ISSN: 1120-4052 printed, 2240-4538 online DOI: 10.7320/FlMedit26.001 Copyright © by International Foundation pro Herbario Mediterraneo, Palermo Contents V. Hugonnot & L. Chavoutier: A modern record of one of the rarest European mosses, Ptychomitrium incurvum (Ptychomitriaceae), in Eastern Pyrenees, France . 5 P. Chène, M. -

2005 Escursioni Con Le Sezioni
to - Taxe Perçue Poste Italiane Spa - Spedizione in Abbonamento Postale D.L. 353/2003 (conv. L. 27/02/2004 N. 46) art. 1, comma 2, DCB Tren EscursioniEscursioni concon lele SezioniSezioni SATSAT -- 20052005 (a cura di Pierandrea Krentzlin) Supplemento al “Bollettino SAT” N. 1 - 2005 Rivista trimestrale 1 Presentazione ’ edizione 2005 della pubblicazione “Escursioni con le Sezioni SAT” costituisce un documento significativo del lavoro generosamente volontario delle Sezio- L ni e dell’impegno dei Soci a partecipare alla vita associativa. Alpinismo, escursionismo, sci-alpinismo, conferenze, attività culturali, incontri, feste e raduni, sono iniziative che vengono proposte con interpretazioni diverse, ma con il comune denominatore di una partecipazione gioiosa, in amicizia, sollecitata da parte di chi ha voglia di stare insieme, condividendo ideali e comportamenti. L’aspetto culturale che spesso caratterizza le varie proposte volte alla conoscenza del mondo della montagna, è un presupposto essenziale per realizzare un giusto approccio ad esso, per comprendere i valori e per salvaguardarli. Interessante è il diffondersi delle gite intersezionali: ciò favorisce il moltiplicarsi delle amicizie e lo scambio di esperien- ze, particolarmente importante ed apprezzato dai giovani. La diversificazione dei pro- grammi, la sapiente distribuzione di gite rese accessibili ai soci con diverso grado di capacità alpinistica o di esperienza culturale, caratterizzano le gite SAT, sempre più gradite ed apprezzate, grazie anche ai nostri bravissimi, esperti accompagnatori. Questo “ben operare” conferma la nostra già vasta rappresentanza che si trasforma in una rappresentatività che si estende ben oltre i confini organizzativi della SAT su- scitando adesione e interesse in ampi strati della società trentina. La SAT dunque si conferma una realtà fortemente radicata, con le sue Sezioni, sul territorio, capace di costruire e offrire un insieme straordinario di proposte culturali, formative e ricreati- ve mai banali o scontate che, in molti casi, raggiungono livelli di assoluta eccellenza. -

Decreti, Delibere E Ordinanze Ministeriali
21-2-2013 GAZZETTA U FFICIALE DELLA R EPUBBLICA I TALIANA Serie generale - n. 44 DECRETI, DELIBERE E ORDINANZE MINISTERIALI MINISTERO DELL’AMBIENTE E DELLA Considerato inoltre che alcuni Stati membri non han- TUTELA DEL TERRITORIO E DEL MARE no proposto siti suf [ cienti a rispondere ai requisiti del- la direttiva 92/43/CEE per certi tipi di habitat e specie, cosicché non si può concludere che la rete Natura 2000 DECRETO 31 gennaio 2013 . sia completa, e che a causa del ritardo nel ricevere le informazioni e nel raggiungere un accordo con gli Stati Sesto elenco aggiornato dei siti di importanza comunita- [ Membri potrà essere necessaria una successiva revisio- ria per la regione biogeogra ca alpina in Italia. ne della lista comunitaria per la regione biogeogra[ ca alpina; IL MINISTRO DELL’AMBIENTE E DELLA Considerato in [ ne che le conoscenze sull’esistenza e TUTELA DEL TERRITORIO E DEL MARE la distribuzione di alcuni habitat naturali dell’allegato I e specie dell’allegato II alla direttiva 92/43/CEE riman- Vista la direttiva 92/43/CEE del Consiglio del 21 mag- gono incomplete per cui in effetti non si può concludere gio 1992, relativa alla conservazione degli habitat naturali se la rete Natura 2000 sia completa o incompleta e si po- e seminaturali e della \ ora e della fauna selvatiche, e in trebbe perciò rendere necessaria una revisione della lista particolare l’art. 4, paragrafo 2, terzo comma; comunitaria per la regione biogeogra [ ca alpina; Visto il decreto del Presidente della Repubblica 8 set- Vista la decisione della Commissione europea tembre 1997, n.