Distribution and Possible Paleoecological Significance Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lithium Concentrations in Brine Springs Near Lake Winnipegosis
Lithium concentrations in brine springs near Lake Winnipego- GS2018-10 sis, west-central Manitoba (parts of NTS 63C, 62N16, 62O12, 13) 1 In Brief: by M.P.B. Nicolas and S.E. Grasby • Brine springs along Lake Winnipegosis contain low levels of lithium, averaging Summary 1263 ppb The most common and cost-effective source of lithium within the brine deposits • Goundwater mixing has diluted the lithium concen- are deep brines in continental sedimentary basins. Southern Manitoba has a complex tration coming out from the springs groundwater aquifer system, with salinities ranging from brines in the deeper aquifers • Deep subsurface Winnipeg and brine springs to freshwater in the shallower and eastern aquifers. The brine springs Formation brines have good potential as a lithium source occur along and near the shores of Lake Winnipegosis on large salt flats and have chemi- cal signatures indicative of mixed halite dissolution brine and freshwater. Lithium concen- trations in these brines range from 150 to 6300 ppb, with an average of 1263 ppb. Citation: Nicolas, M.P.B. and Grasby, S.E. 2018: Lithium concentra- Introduction tions in brine springs near Lake Saline groundwater in continental sedimentary basins are generated from evaporitic Winnipegosis, west-central Manitoba (parts of NTS 63C, concentration and/or halite dissolution, where the former is a primary process and the 62N16, 62O12, 13); in Report latter is a secondary (diagenetic) process (Clayton et al., 1966; Walter et al., 1990). Both of Activities 2018, Manitoba these mechanisms provide opportunities for trace elements to accumulate, potentially Growth, Enterprise and Trade, Manitoba Geological Survey, reaching economic concentrations. -

Faculty of Graduate Studies
3'A.C|IJL:IY 0F t'RA:)LlrÊ'TE Ë'TJIÏE'1 DEP/ìRTMfiÌT OF,- T],VJ ]. EI$GTIVEEBSNI] tilIÏVElRlrllty 03' j4AN:]"T0BA AN ÍN\TfißTTI.íTITTOJ\T O}' T}IE ÐTVERåJ.OIf C}:.rÎ NOR'IIìTB.N }4ANTTOEA I{ATE¡I.:3 J,NTO ].,AKE }IIAJü OÉA l- Jt t. .4. Fl:-¡40I{ n TI]Tfl:I:ì J:I{,PÅR]TÏAL FLI"LF],ffÆI\T 'sUBT\MTTEÐ CIF T'TIE REQLÍSAEME}Í?i3 iIÜR TTIE NEGBEE ÐF j/ÛslER Olr SÜ3,Ï:IùGE APRTT L?6"r acitro\'fT.m)Gm{msrs The ç'råter v¡Èshes to e:qpress hi s mrnest appreciatf on to Jan:ice, A.LLísou and. Davåd., Írz acknowledgenaent of thefr patÍenee anð u.nåerstaadågg ín vi.ll-iragly glvfrug 'ap thefr ri.ghts to 'the eompanS"orashåp anå at'Lentínn r¡f a husba.trð and' fether during the many hours sPent on this thesis. I&ny that:les ave alnc¡ åt¿Ë to Professor E" Ktliper for his årrspåratiou in the i,nitåatlon of, this stud-y and' his guldance ancl assåstanee tÏ.lz"oughout " The w.riter would also Like to eryrers his appreciation to the månage¡geni and ,sf"aff af Underwooð, Mçtellan and. AssocÍates, for their ånteres'L a.rio Ínvolvement ån thj"s thesis, especially to Meser"s" A. G. Spaffor"d" and R. c. La'tta, vå'bhout whose gtrÍdance and assÍstance this thesís j..n j"ts fånãt fow¡r wou[d. t ot be possfble' The r+¡"íter also wÍshes to acknovf-ed-ge grateful-Ly the assfs.tance of 3,fr. -
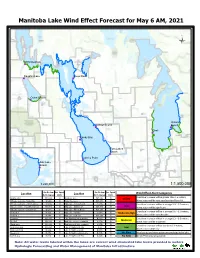
Manitoba Lake Wind Effect Forecast for May 6 AM, 2021 Ü
Manitoba Lake Wind Effect Forecast for May 6 AM, 2021 Ü Winnipegosis !P Dauphin Lake Steep Rock !P !P Ochre Beach !P Victoria Lundar Beach Beach !P !P Gimli !P Sandy Bay !P Twin Lakes Beach !P Lynchs Point !P Oak Lake Beach !P 1:220,000 1:1,500,000 Ave.Dir.from Ave. Speed Ave.Dir.from Ave. Speed Wind Effect Alert Categories Location North (degree) (km/h) Location North (degree) (km/h) Crest level increase will be greater than 1.5 meters; Dauphin Lake NW (320) 21 Manipogo area NW (336) 15 Severe MB Lake west side - Sandy Bay W (292) 18 Crane River area NW (335) 12 Wave action will be very significant and forceful. MB Lake south side - Twin Beaches W (274) 17 Wpg Lake - Victoria Beach W (289) 12 Crest level increase will be in a range 0.9 ~ 1.5 meters; High MB Lake middle - Vogar/The Narrows NW (311) 12 Wpg Lake - Gimli Beach NW (318) 13 Wave action will be significant. MB Lake east side - Lundar Beach W (281) 12 Wpg Lake - Alberta Beach NW (293) 11 Crest level increase will be in a range 0.6 ~ 0.9 meters; Northend 1 N (351) 9 Wpg Lake - O'Hanly W (292) 11 Moderate-High Northend 2 N (344) 11 Wpg Lake south side NW (294) 11 Wave action will be considerable. Guynemer NW (337) 12 Lake Winnipegosis south side N (343) 19 Crest level increase will be in a range 0.3 ~ 0.6 meters; Moderate The Bluff NW (331) 14 Oak Lake east side community W (276) 18 Wave action will be moderate. -

CHARACTERISTICS and DISTRIBUTION of LAKES 631 Lake Winnipegosis, Lying to the North of Lake Manitoba
CHARACTERISTICS AND DISTRIBUTION OF LAKES 631 Lake Winnipegosis, lying to the north of Lake Manitoba. at an elevation of 828 feet above sea-level, is about 130 miles long, by 20 miles in maximum breadth. It covers an area of 2080 square miles, and has a maximum depth of 38 feet. It is fed by many small streams from the west, and by the overflow of Lake Dauphin (840 feet above sea-level) through Mossy River. The outlet is by the very indirect way of Waterhen River, through Waterhen Lake, to Lake Manitoba. The total area of the lakes in the Winnipeg basin is 13,500 square miles. Lake Wollaston, 1300 feet above sea-level, is the ultimate source River of the Reindeer River, one of the chief tributaries of the Churchill Churchill. River. It is about 800 square miles in area, and discharges by two outlets—to the north by the Stone River into the extreme eastern arm of Lake Athabasca, and to the south-east by the Cochrane River into Reindeer Lake. Reindeer Lake, 1150 feet above sea-level, is 135 miles long, and has an area of 2490 square miles; the Reindeer River carries its overflow to the Churchill River. The rivers in Labrador are often like strings of lakes, and divide River Rupert. and unite again in their course, while the lakes frequently discharge in two directions. Lake Mistassini, 1350 feet above sea-level, the largest lake, is practically two parallel lakes divided by a range of islands in the centre. The western lake is 90 miles long by 13 to 17 miles wide, and the eastern lake is 60 miles long by 5 to 10 miles wide, the greatest depths being 300 to 400 feet. -

RIVERS and LAKES 11 the Great Lakes.—Table 4 Shows the Length
RIVERS AND LAKES 11 The Great Lakes.—Table 4 shows the length, breadth, area, elevation above sea-level and maximum depth of each of the Great Lakes. i.—Area, Elevation and Depth of the Great Lakes. Maximum Elevation Lakes. Length. Breadth. depth. Area. above sea-level. square miles. miles. feet. miles. feet. Superior... 383 160 1,180 31,810 602-29 Michigan.. 320 118 870 22,400 581-13 Huron 247 101 750 23,010 581-13 St. Clair.., 26 24 23 460 575-62 Erie 241 57 210 9,940 572-52 Ontario 180 53 738 7,540 246-17 Lake Superior, with its area of 31,810 square miles, is the largest body of fresh water in the world. As the international boundary between Canada and the United States passes through the centre of lakes Superior, Huron, Erie, St. Clair and Ontario, only half of the areas of these lakes given in the above statement is Canadian. The whole of lake Michigan is within United States territory. From the western end of lake Superior to the mouth of the St. Lawrence there is, with the aid of the canal system, a continuous navigable waterway. The total length of the St. Law rence river from the head of the St. Louis river to Pointe-des-Monts, at the entrance of the gulf of St. Lawrence, is 1,900 miles. The tributaries of the St. Lawrence, several of which have themselves important tributaries, include the Ottawa river, 685 miles long, the St. Maurice river, 325 miles long, and the Sague- nay (to head of Peribonka), 405 miles long. -

05260 Sask Watershed Report.Qxd:State of The
Saskatchewan Watershed Authority STATE OF THE WATERSHED REPORTING FRAMEWORK Monitoring and Assessment Branch Stewardship Division January 2006 Suite 420-2365 Albert Street Regina, Saskatchewan S4P 4K1 www.swa.ca Cover photos courtesy of (top-bottom): Saskatoon Tourism; Nature Saskatchewan; Saskatchewan Watershed Authority; Saskatchewan Watershed Authority; Wawryk Associates Ltd.; Ducks Unlimited Canada; Saskatchewan Watershed Authority; Nature Saskatchewan; and Saskatchewan Industry and Resources. Minister’s Message From the grasslands in the southwest to the Taiga Shield lakes of the north, we in Saskatchewan are blessed with richly diverse ecosystems. Over the past few decades, we have come to understand more about the nature of these ecosystems, and how we rely on them to support our economy and our overall quality of life. We have also increased our understanding of the responsibility we bear to assess, protect and improve our province’s environmental health, in balance with our social and economic priorities, for the good of present and future generations. We face a number of challenges regarding our water’s quality and availability. It is up to Saskatchewan people to determine how we will meet these challenges. The Saskatchewan government has taken a leadership role in assessing, protecting and managing our source waters by addressing them at the watershed level. The State of the Watershed Reporting Framework represents a critical step in understanding our relationship with Saskatchewan’s watersheds, as well as our role as stewards of our environment. For the first time in Saskatchewan, this framework will integrate the information collected by numerous provincial and federal agencies and present it in an easily understandable, technically sound report- card format. -

MANITOBA's ECOCLIMATIC REGIONS Geoffrey A.J. Scott
MANITOBA'S ECOCLIMATIC REGIONS Geoffrey A.J. Scott 4 Although humans have an enor- matic provinces, and portions of 4 of etation zones and soil types. mous potential to modify natural the 10 Canadian ecoclimatic prov- It is the overall role of climate, ecosystems, many human activities inces — the Grassland, Boreal, however, that accounts for the are both stimulated and/or limited Subarctic, and Arctic — are found northwest-to-southeast orientation by constraints imposed by the eco- in Manitoba (Figure 4.1).3 Because of Manitoba's major ecoclimatic re- system or climate. Whereas much of of their large size and internal vari- gions and zonal vegetation and southern and west-central Mani- ation, these provinces are subdi- soils. Types of vegetation cover and toba has already been drastically vided into ecoclimatic regions, and the distribution of freely drained modified during the last century by 8 such regions are represented in soils correlate strongly with in- agriculture, grazing, forestry, and Manitoba. creasing moisture availability as urban development to the point As the definition implies, each precipitation increases from west to that its original natural-vegetation ecoclimatic region has its own char- east (Figure 4.4), and also with in- covers are hardly recognizable, acteristic combination of soil types, creasing effective precipitation as many other regions in the north and vegetation cover, and wildlife that one moves from south to north. northeast appear quite pristine. An makes it distinct from any other re- Wetlands, however, -

LAKES 15 the Great Lakes.—Table 4 Shows the Length, Breadth, Area
LAKES 15 The Great Lakes.—Table 4 shows the length, breadth, area, elevation above sea-level and maximum depth of each of the Great Lakes. Particularly notable is the depth of lake Superior and the shallowness of lake St. Clair and lake Erie. i.—Area, Elevation and Depth of the Great Lakes. Elevation Lake. Length. Breadth. Maximum Area. above Depth. Sea-level. miles. miles. feet. square feet. miles. 383 160 1,180 31,810 602-29 320 118 870 22,400 581-13 Huron 247 101 750 23,010 581-13 St. Clair 26 24 23 460 575-62 Erie 241 57 210 9,940 572-52 180 53 738 7,540 246-17 Lake Superior, with its area of 31,810 square miles, is the largest body of fresh water in the world. As the International Boundary between Canada and the United States passes through the centre of lakes Superior, Huron, Erie, St. Clair and Ontario; only a part of the areas of these lakes given in the above statement is Canadian. The whole of lake Michigan is within United States territory. From the western end of lake Superior to the mouth of the St. Lawrence there is, with the aid of the canal system, a continuous navigable waterway. The total length of the St. Lawrence waterway from the head of the St. Louis river in Minnesota to Pointe-des-Monts, at the entrance of the gulf of St. Lawrence, is 1,900 miles. The tributaries of the St. Lawrence, several of which have themselves important tributaries, include the Ottawa river, 685 miles long, the St. -

Appendix 1. Details Regarding Historical and Current Colonial Waterbird Surveys on the Large Lakes of Southern Manitoba, Canada
Appendix 1. Details regarding historical and current colonial waterbird surveys on the large lakes of southern Manitoba, Canada. Table A1.1. Colonial waterbird surveys conducted between 1969 and 2017 in southern Manitoba. Details include survey dates when available, the portion of the region surveyed (where ‘Whole region’ = Lake Winnipeg, Lake Winnipegosis, Lake Manitoba, and the Interlake region), survey method when available (where ‘Aerial’ = fixed wing, helicopter, or a combination; ‘Boat’ = counts made from a boat; ‘Ground’ = counts made within the colony; or a combination), and species included in the survey. Year Dates Area surveyed Survey method Species Reference 1969 “when the eggs Whole region Aerial (visual)/ DCCO, Vermeer 1969, 1970a started to hatch” Ground AWPE 1970 24 June Lake Winnipegosis† Ground CATE Vermeer 1970b 1979 11, 12, 19 June Whole region Aerial (visual)/ All species Koonz and Rakowski Ground 1985 1986 NA Whole region NA CATE B. Koonz, cited in Wires and Cuthbert 2000 1987 Late June Lake Winnipegosis Boat/Ground DCCO Hobson et al. 1989 1989-92 NA Whole region NA CATE B. Koonz, cited in Wires and Cuthbert 2000 1990 NA Lake Winnipeg, Lake NA COTE Manitoba Conservation, Manitoba, Inter-lake as cited in Wilson et al. region 2014 1999 4-11 June Lake Winnipegosis Boat/Ground All species Koonz 2000 2012 13, 15, 16 June Whole region Aerial (photography) All species Wilson et al. 2014 2017 11, 12, 19, 20 June Whole region Aerial (photography) All species Current study †Surveys were also conducted at several colonies on Lake Winnipeg, but these were not included in our analysis LITERATURE CITED Hobson, K. -

Lake Manitoba Water Qality & Ecosystem Health Workshop
Lake Manitoba and its Watershed: Knowledge Gaps & Next Steps Sunrise over Lake Manitoba at Delta Marsh (© Marliese Peterson) TABLE OF CONTENTS 1. Executive Summary ................................................................................................................. 1 2. Introduction.............................................................................................................................. 2 2.1. Background to Lake Manitoba .......................................................................................... 2 2.2. Rationale for workshop ..................................................................................................... 2 3. Water Levels & Regulation..................................................................................................... 4 3.1. Presentations ..................................................................................................................... 4 3.2. Knowledge Gaps ............................................................................................................... 4 3.1. Next Steps For Top-Ranked Knowledge Gaps ................................................................. 8 4. Watershed Management & Landscape Processes .............................................................. 12 4.1. Presentations ................................................................................................................... 12 4.2. Knowledge Gaps ............................................................................................................ -

An Annotated Bibliography on Lake Manitoba and Adjoining Waters
Delta Marsh Field Station (University of Manitoba) Occasional Publication No. 3 AnAn AnnotatedAnnotated BibliographyBibliography onon LakeLake ManitobaManitoba andand AdjoiningAdjoining WatersWaters byby TaraTara L.L. BortoluzziBortoluzzi Delta Marsh Field Station (University of Manitoba) Box 38, Rural Route 2 Portage la Prairie, MB, Canada R1N 3A2 Telephone: (204) 857-8637 Fax: (204) 857-4683 E-mail: [email protected] Web: www.umanitoba.ca/delta_marsh Delta Marsh Field Station (University of Manitoba) Occasional Publication, Number 3 (November 2003) Edited by L. Gordon Goldsborough Delta Marsh Field Station (University of Manitoba) ISSN 1491-5154 ISBN #-#######-#-# © 2003, Delta Marsh Field Station (University of Manitoba) The cover photo shows the sun setting over Lake Manitoba, 27 July 2004, with bulrushes on the beach by the Delta Marsh Field Station in the foreground. Photo: Heidi den Haan. Copies of this report are available on the Internet at: www.umanitoba.ca/delta_marsh/pubs Suggested bibliographic format for this report: Bortoluzzi, T. L. 2003. An Annotated Bibliography on Lake Manitoba and Adjoining Waters. Delta Marsh Field Station (University of Manitoba) Occasional Publication No. 3, Winnipeg, Canada. ## pp. An Annotated Bibliography on Lake Manitoba and Adjoining Waters by Tara L. Bortoluzzi Department of Botany, University of Manitoba, Winnipeg, Manitoba, Canada Delta Marsh Field Station (University of Manitoba) Occasional Publication No. 3 November 2003 SUMMARY Keywords: TEXT. An Annotated Bibliography on Lake Manitoba and Adjoining Waters by Tara L. Bortoluzzi Department of Botany, University of Manitoba, Winnipeg, Canada Delta Marsh Field Station (University of Manitoba) Occasional Publication No. 3 November 2003 Delta Marsh Field Station (University of Manitoba) 239 Machray Hall Winnipeg, MB, Canada R3T 2N2 Telephone: (204) 474-9297 Fax: (204) 474-7618 E-mail: [email protected] Web: www.umanitoba.ca/delta_marsh Delta Marsh Field Station (University of Manitoba) Occasional Publication, Number 3 (November 2003) Edited by L. -

WATERHEN LAKE (Inc: Chitek, Inland, Archie & Crab) Summary Report
Wildlife & Fisheries Branch Report 2017 – 02 Waterhen Lake Summary Report (Inc: Chitek, Inland, Archies & Crab) William Galbraith, Geoff Klein & Gord Kirbyson Manitoba Sustainable Development 2017 Table of Contents Page Location ……...................................................................................................... 3 Communities ….................................................................................................... 4 Fish Species ......................................................................................................................... 4 Types of Fisheries ……......................................................................................................... 4 1. Aboriginal Domestic/Sustenance Fishing ........................................................................... 4 2. Commercial Net ................................................................................................................... 4 3. Commercial Tourism ……..................................................................................................... 5 4. Recreational Angling ……..................................................................................................... 5 History of the Fishery …….................................................................................... 5 1. Commercial ........................................................................................................................ 5 2. Recreational .......................................................................................................................