Acid Citrus Fruit Improvement Via Interploid Hybridization Using
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lulav-And-Etrog-Instructions.Pdf
אֶּתְ רֹוג לּולָב LULAV AND ETROG: THE FOUR SPECIES What they are and what to do with them INTRODUCTION The commandment regarding the four species (of the lulav and etrog) is found in the Torah. After discussing the week-long Sukkot festival, specific instructions for how to celebrate the holiday are given. Leviticus 23:40 instructs: םּולְקַחְתֶּ לָכֶּם בַּיֹוםהָרִ אׁשֹון פְרִ י עֵץ הָדָרכַפֹּת תְ מָרִ ים וַעֲנַף עֵץ־עָבֹּת וְעַרְ בֵי־נָחַל ּושְ מַחְתֶּ ם לִפְ נֵי ה' אֱֹלהֵיכֶּם ׁשִבְ עַת יָמִ ים “On the first day you shall take the product of hadar trees, branches of palm trees, boughs of leafy trees, and willows of the brook, and you shall rejoice before Adonai your God seven days." These are the four species that form the lulav and etrog. The four species are waved in the synagogue as part of the service during the holiday of Sukkot. Traditionally, they are not waved on Shabbat because bringing these items to the synagogue would violate the prohibition against carrying. Some liberal synagogues do wave the lulav and etrog on Shabbat. While it is customary for each individual to have a lulav and etrog, many synagogues leave some sets in the synagogue sukkah for the use of their members. The lulav and etrog may also be waved at home. Below you will find some basic information about the lulav and etrog, reprinted with permission from The Jewish Catalogue: A Do-It-Yourself Kit, edited by Richard Siegel, Michael Strassfeld and Sharon Strassfeld, published by the Jewish Publication Society. HOW THE FOUR PARTS FIT TOGETHER The lulav is a single palm branch and occupies the central position in the grouping. -

New Acid Citrus Selections for Florida
New acid citrus selections for Florida Lemon and lime-like selections with niche market potential are being developed with biotechnology at the University of Florida. By Jude Grosser, Zenaida Viloria and Manjul Dutt re you ready for a purple margarita? Would you like a fragrant, juicy lime is a naturally occurring citrus lemon for your iced tea with no seeds to clog your straw or dishwasher triploid, which is why it is seedless. drain? How about some seedless lime trees that are cold-hardy enough The new seedless watermelons in the Afor Central Florida? These and many more interesting acid-citrus marketplace are also triploids. selections are now on the horizon, including some with good ornamental potential. Due to the fact that new and This article will focus on progress in using emerging biotechnologies to develop improved citrus cultivars must be interesting new citrus cultivars in the lemon and lime group. Cultivars include seedless to compete in the national some that will not have regulatory constraints, and also a genetically modified and international marketplace, the organism (GMO)-derived purple Key lime as a teaser for the future. University of Florida’s Citrus Research and Education Center (UF/CREC) LEARNING FROM they are triploids. People and most citrus improvement team (working THE BANANA citrus trees are diploid, meaning with Fred Gmitter) has formulated Have you ever wondered why you there are two sets of chromosomes in several ways to create triploids as a key never find seeds in your bananas? Did each cell. Triploid bananas have three method of developing seedless citrus you know that there are wild-type sets of chromosomes per cell. -

"Performance of Citrus Scion Cultivars and Rootstocks in a High-Density
REPORTS HORTSCIENCE 26(7):837-840. 1991. house and planted in the field in 1981. A split plot experiment and analysis of variance Performance of Citrus Scion Cultivars (ANOVA) statistics were used with four rep- lications, with cultivar as the main plot and and Rootstock in a High-density rootstock as the subplot. Field plots were four ´ four trees, with data taken from the Planting center four of the 16 trees. They were planted 1.5 m in the row and 3.3 m between rows T.A. Wheaton, W.S. Castle, J.D. Whitney, and D.P.H. Tucker and were irrigated and fertigated as required Citrus Research and Education Center, University of Florida, Institute of to maintain optimal soil water and nutrient levels using one microsprinkler per two trees. Food and Agricultural Sciences, 700 Experiment Station Road, Lake Trees were mechanically hedged and topped Alfred, FL 33850 during Summer 1987 and hedged again in 1989 to maintain a 1.5-m alley between rows Additional index words. tree spacing, yield efficiency and a 2.5-m tree height. Thus, the canopy Abstract. ‘Hamlin’ and ‘Valencia’ oranges [Citrus sinensis (L.) Osb.], ‘Murcott’ tangor size allocated for each tree was 1.5 m in the (C. reticulata Blanco ´ C. sinensis), and ‘Redblush’ grapefruit (C. paradisi Macf.) on row, 1.8 m across the row, and 2.5 m in 15 rootstock and own-rooted cuttings were planted at a 1.5 ´ 3.3-m spacing providing height, providing 6.8 m3 of canopy volume a density of 2020 trees/ha. -

Bright Citron™ Soak
Product Profile BRIGHT CITRON™ SOAK WHAT IT IS An aromatic and refreshing sea salt soak for cleansing skin. WHAT IT DOES Softens and cleanses skin. WHY YOU NEED IT • Provides the first step to cleanse impurities and soften skin for a luxurious manicure or pedicure experience. • Gently cleanses without drying skin. FRAGRANCE FEATURED Pink Grapefruit and Warm Amber. FRAGRANCE PROFILE • Bright citrus top notes. • Soft floral mid notes. CREATIVE SUGGESTIONS • Light woody base notes. • Create a fragrant ceremonial experience by placing the Soak into warm water directly prior to ACTIVE BOTANICALS immersing hands or feet. Kaffir Lime (Citrus Hystrix Leaf Extract): • Add lime or grapefruit slices to enhance the Is known to purify skin and promote radiance. experience. • Use as a foot soak prior to all salon services. Honey: • Excellent retail product for home use in bath tubs. Is known to hydrate skin and help retain moisture. INGREDIENT LISTING FEATURED INGREDIENTS & BENEFITS Maris Sal ((Sea Salt) Sel Marin), Sodium Sesquicarbonate, Sea Salt: Argania Spinosa Kernel Oil, Honey, Citrus Hystrix Leaf Natural salt used for its therapeutic properties. Extract, Aqua ((Water) Eau), Glycerin, Propylene Glycol, Salts increase the flow of water in and out of Hydrated Silica, Parfum (Fragrance), Limonene, Linalool, cells, in essence flushing and cleansing the cells. Hexyl Cinnamal, Citral. This facilitates the absorption of other ingredients into the cells. AVAILABLE SIZES • Retail/Professional: 410 g (14.4 oz) Argan Oil (Argania Spinosa Kernel Oil): • Professional: 3.3 kg (118.8 oz) Is known to nourish dry skin. DIRECTIONS FOR USE • For feet, add a teaspoon to footbath and agitate water to dissolve. -

Known Host Plants of Huanglongbing (HLB) and Asian Citrus Psyllid
Known Host Plants of Huanglongbing (HLB) and Asian Citrus Psyllid Diaphorina Liberibacter citri Plant Name asiaticus Citrus Huanglongbing Psyllid Aegle marmelos (L.) Corr. Serr.: bael, Bengal quince, golden apple, bela, milva X Aeglopsis chevalieri Swingle: Chevalier’s aeglopsis X X Afraegle gabonensis (Swingle) Engl.: Gabon powder-flask X Afraegle paniculata (Schum.) Engl.: Nigerian powder- flask X Atalantia missionis (Wall. ex Wight) Oliv.: see Pamburus missionis X X Atalantia monophylla (L.) Corr.: Indian atalantia X Balsamocitrus dawei Stapf: Uganda powder- flask X X Burkillanthus malaccensis (Ridl.) Swingle: Malay ghost-lime X Calodendrum capense Thunb.: Cape chestnut X × Citroncirus webberi J. Ingram & H. E. Moore: citrange X Citropsis gilletiana Swingle & M. Kellerman: Gillet’s cherry-orange X Citropsis schweinfurthii (Engl.) Swingle & Kellerm.: African cherry- orange X Citrus amblycarpa (Hassk.) Ochse: djerook leemo, djeruk-limau X Citrus aurantiifolia (Christm.) Swingle: lime, Key lime, Persian lime, lima, limón agrio, limón ceutí, lima mejicana, limero X X Citrus aurantium L.: sour orange, Seville orange, bigarde, marmalade orange, naranja agria, naranja amarga X Citrus depressa Hayata: shiikuwasha, shekwasha, sequasse X Citrus grandis (L.) Osbeck: see Citrus maxima X Citrus hassaku hort. ex Tanaka: hassaku orange X Citrus hystrix DC.: Mauritius papeda, Kaffir lime X X Citrus ichangensis Swingle: Ichang papeda X Citrus jambhiri Lushington: rough lemon, jambhiri-orange, limón rugoso, rugoso X X Citrus junos Sieb. ex Tanaka: xiang -

Tropical Horticulture: Lecture 32 1
Tropical Horticulture: Lecture 32 Lecture 32 Citrus Citrus: Citrus spp., Rutaceae Citrus are subtropical, evergreen plants originating in southeast Asia and the Malay archipelago but the precise origins are obscure. There are about 1600 species in the subfamily Aurantioideae. The tribe Citreae has 13 genera, most of which are graft and cross compatible with the genus Citrus. There are some tropical species (pomelo). All Citrus combined are the most important fruit crop next to grape. 1 Tropical Horticulture: Lecture 32 The common features are a superior ovary on a raised disc, transparent (pellucid) dots on leaves, and the presence of aromatic oils in leaves and fruits. Citrus has increased in importance in the United States with the development of frozen concentrate which is much superior to canned citrus juice. Per-capita consumption in the US is extremely high. Citrus mitis (calamondin), a miniature orange, is widely grown as an ornamental house pot plant. History Citrus is first mentioned in Chinese literature in 2200 BCE. First citrus in Europe seems to have been the citron, a fruit which has religious significance in Jewish festivals. Mentioned in 310 BCE by Theophrastus. Lemons and limes and sour orange may have been mutations of the citron. The Romans grew sour orange and lemons in 50–100 CE; the first mention of sweet orange in Europe was made in 1400. Columbus brought citrus on his second voyage in 1493 and the first plantation started in Haiti. In 1565 the first citrus was brought to the US in Saint Augustine. 2 Tropical Horticulture: Lecture 32 Taxonomy Citrus classification based on morphology of mature fruit (e.g. -

Marketing Standards for Citrus Fruits
COMMISSION IMPLEMENTING REGULATION (EU) No 543/2011 of 7 June 2011 laying down detailed rules for the application of Council Regulation (EC) No 1234/2007 in respect of the fruit and vegetables and processed fruit and vegetables sectors. CONSOLIDATED TEXT: Annex I; Part 2 of Part B MARKETING STANDARD FOR CITRUS FRUIT I. DEFINITION OF PRODUCE This standard applies to citrus fruit of varieties (cultivars) grown from the following species, to be supplied fresh to the consumer, citrus fruit for industrial processing being excluded: - lemons grown from the species Citrus limon (L.) Burm. f. and hybrids thereof, - mandarins grown from the species Citrus reticulata Blanco, including satsumas (Citrus unshiu Marcow), clementines (Citrus clementina hort. ex Tanaka), common mandarins (Citrus deliciosa Ten.) and tangerines (Citrus tangerina Tanaka) grown from these species and hybrids thereof, - oranges grown from the species Citrus sinensis (L.) Osbeck and hybrids thereof. II. PROVISIONS CONCERNING QUALITY The purpose of the standard is to define the quality requirements for citrus fruit after preparation and packaging. However, at stages following dispatch products may show in relation to the requirements of the standard: - a slight lack of freshness and turgidity, - for products graded in classes other than the “Extra” Class, a slight deterioration due to their development and their tendency to perish. A. Minimum requirements In all classes, subject to the special provisions for each class and the tolerances allowed, the citrus fruit must be: - intact, - free of bruising and/or extensive healed overcuts, CI.1 - sound; produce affected by rotting or deterioration such as to make it unfit for consumption is excluded, - clean, practically free of any visible foreign matter, - practically free from pests, - free from damage caused by pests affecting the flesh, - free of signs of shrivelling and dehydration, - free of damage caused by low temperature or frost, - free of abnormal external moisture, - free of any foreign smell and/or taste. -

Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Relatives
Chemical variability of peel and leaf essential oils in the Citrus subgenus Papeda (Swingle) and relatives Clémentine Baccati, Marc Gibernau, Mathieu Paoli, Patrick Ollitrault, Félix Tomi, François Luro To cite this version: Clémentine Baccati, Marc Gibernau, Mathieu Paoli, Patrick Ollitrault, Félix Tomi, et al.. Chemical variability of peel and leaf essential oils in the Citrus subgenus Papeda (Swingle) and relatives. Plants, MDPI, 2021, 10 (6), pp.1117. 10.3390/plants10061117. hal-03262123 HAL Id: hal-03262123 https://hal.archives-ouvertes.fr/hal-03262123 Submitted on 16 Jun 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution| 4.0 International License Chemical variability of peel and leaf essential oils in the Citrus subgenus Papeda (Swingle) and relatives Clémentine Baccati 1, Marc Gibernau 1, Mathieu Paoli 1, Patrick Ollitrault 2,3, Félix Tomi 1, * and François Luro 2 1 Université de Corse-CNRS, UMR 6134 SPE, Route des Sanguinaires, 20000 Ajaccio, France; [email protected] (C.B.) ; [email protected] (M.G.) ; [email protected] (M.P.) ; [email protected] (F.T.) 2 UMR AGAP Institut, Univ Montpellier, CIRAD, INRAE, Institut Agro – 20230, San Giuliano, France 3 CIRAD, UMR AGAP, F-20230 San Giuliano, France * Correspondence: [email protected]; tel.:+33-495-52-4122. -

Crop Profile for Citrus (Minor) Lime, Pummelo, and Kumquat in Florida
Crop Profile for Citrus (Minor) Lime, Pummelo, and Kumquat in Florida Prepared: November, 2001 General Production Information ● Lime in this crop profile refers to the 'Tahiti' lime (Citrus latifolia). Pest management practices of other members of the Rutaceae such as Key lime (Citrus aurantifolia), pummelo (Citrus grandis), and kumquat (Fortunella sp.) may also be described, since survey questionnaires grouped these four tropical fruits together. All of these species of citrus are grown for the fresh market. ● The 1997-1998 average yield of limes in Florida was 14,344 pounds per acre. At a price of $5.02 per box, the Florida crop (440,000 boxes) was worth approximately $2.21 million (1). ● In 2000-2001, 1,171 acres in Florida produced 22 million pounds of 'Tahiti' lime. This is a 58 percent decrease from the previous year, when 53 million pounds were recorded (2,3). ● As of the end of 2000, there has been a 63 percent reduction in lime acreage since 1996 (bearing acreage of 3,168 acres). Except for 20 acres, all of the lime production is located in Miami-Dade County. Consequently, ninety-eight percent of 'Tahiti' lime production occurs in this county (3). ● Florida was ranked first in the United States in lime production. No other states were listed as leading lime-producers (1). ● Reductions in lime acreage have occurred due to Hurricane Andrew and citrus canker. Bearing acreage before Andrew was approximately 6,000 acres, which dropped drastically to 1,668 acres after this event (4). An estimated 290,000 lime trees (2,800 acres) have been lost to citrus canker in the 2000-2001 period (2). -

About Limes by George Geary CCP
All About Limes By George Geary CCP Lime History In the eighteenth century, Scottish naval surgeon Sir James Lind learned by his observation of long-haul sailors that citrus fruits conquered the dreaded scurvy (lack of Vitamin C) which had divested the ranks of the British navy more than any enemy. Between 1795 and 1815, some 1.6 million gallons of lime Juice drastically reduced the mortality rate of seamen. Along with their daily ration of rum, British sailors were required to consume a daily ration of lime Juice; hence British seamen became known as limeys. Since Britain was often at war with Mediterranean countries that exported lemons, limes imported cheaply from the English colony of Jamaica were substituted as the citrus of choice. Key Lime (also known as Mexican Lime and West Indies Lime) Cultivated for thousands of years in the Indo-Malayan region, this variety has long been treasured for its fruit and decorative foliage. The Key lime made its way to North Africa and the Near East via Arabian traders, and then carried on to Palestine and Mediterranean Europe by the Crusaders. Columbus is credited with bringing the Key lime to Haiti, where Spanish settlers to Florida carried it on. It flourished in South Florida, particularly the Florida Keys, hence the current common name of Key lime. Due to hurricane-depleted soils, locals switched from pineapple commercial crops to limes in 1906, and business boomed until a hurricane once again reared and wiped out the lime groves, never to be restored. Today, most Key limes come from Mexico. -
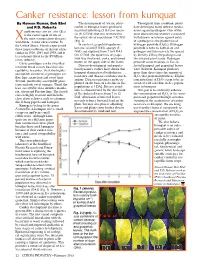
Canker Resistance: Lesson from Kumquat by Naveen Kumar, Bob Ebel the Development of Asiatic Citrus Throughout Their Evolution, Plants and P.D
Canker resistance: lesson from kumquat By Naveen Kumar, Bob Ebel The development of Asiatic citrus Throughout their evolution, plants and P.D. Roberts canker in kumquat leaves produced have developed many defense mecha- anthomonas citri pv. citri (Xcc) localized yellowing (5 DAI) or necro- nisms against pathogens. One of the is the causal agent of one of sis (9-12 DAI) that was restricted to most characteristic features associated the most serious citrus diseases the actual site of inoculation 7-12 DAI with disease resistance against entry X (Fig. 2). of a pathogen is the production of worldwide, Asiatic citrus canker. In the United States, Florida experienced In contrast, grapefruit epidermis hydrogen peroxide (H2O2). Hydrogen three major outbreaks of Asiatic citrus became raised (5 DAI), spongy (5 peroxide is toxic to both plant and canker in 1910, 1984 and 1995, and it DAI) and ruptured from 7 to 8 DAI. pathogen and thus restricts the spread is a constant threat to the $9 billion On 12 DAI, the epidermis of grape- by directly killing the pathogen and citrus industry. fruit was thickened, corky, and turned the infected plant tissue. Hydrogen Citrus genotypes can be classified brown on the upper side of the leaves. peroxide concentrations in Xcc-in- into four broad classes based on sus- Disease development and popula- fected kumquat and grapefruit leaves ceptibility to canker. First, the highly- tion dynamics studies have shown that were different. Kumquat produces susceptible commercial genotypes are kumquat demonstrated both disease more than three times the amount of Key lime, grapefruit and sweet lime. -

048 (3) Plenodomus Tracheiphilus (Formerly Phoma Tracheiphila)
Bulletin OEPP/EPPO Bulletin (2015) 45 (2), 183–192 ISSN 0250-8052. DOI: 10.1111/epp.12218 European and Mediterranean Plant Protection Organization Organisation Europe´enne et Me´diterrane´enne pour la Protection des Plantes PM 7/048 (3) Diagnostics Diagnostic PM 7/048 (3) Plenodomus tracheiphilus (formerly Phoma tracheiphila) Specific scope Specific approval and amendment This Standard describes a diagnostic protocol for First approved in 2004–09. Plenodomus tracheiphilus (formerly Phoma tracheiphila).1 Revision approved in 2007–09 and 2015–04. Phytosanitary categorization: EPPO A2 list N°287; EU 1. Introduction Annex designation II/A2. Plenodomus tracheiphilus is a mitosporic fungus causing a destructive vascular disease of citrus named ‘mal secco’. 3. Detection The name of the disease was taken from the Italian words ‘male’ = disease and ‘secco’ = dry. The disease first 3.1 Symptoms appeared on the island of Chios in Greece in 1889, but the causal organism was not determined until 1929. Symptoms appear in spring as leaf and shoot chlorosis fol- The principal host species is lemon (Citrus limon), but lowed by a dieback of twigs and branches (Fig. 2A). On the the fungus has also been reported on many other citrus spe- affected twigs, immersed, flask-shaped or globose pycnidia cies, including those in the genera Citrus, Fortunella, appear as black points within lead-grey or ash-grey areas Poncirus and Severina; and on their interspecific and (Fig. 3B). On fruits, browning of vascular bundles can be intergenic hybrids (EPPO/CABI, 1997 – Migheli et al., observed in the area of insertion of the peduncle. 2009).