Cryptogams of the Cedar River Watershed September 2001
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
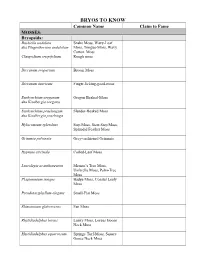
Bryo's to Know Table
BRYOS TO KNOW Common Name Claim to Fame MOSSES: Bryopsida: Buckiella undulata Snake Moss, Wavy-Leaf aka Plagiothecium undulatum Moss, Tongue-Moss, Wavy Cotton, Moss Claopodium crispifolium Rough moss Dicranum scoparium Broom Moss Dicranum tauricum Finger-licking-good-moss Eurhynchium oreganum Oregon Beaked-Moss aka Kindbergia oregana Eurhynchium praelongum Slender-Beaked Moss aka Kindbergia praelonga Hylocomium splendens Step Moss, Stair-Step Moss, Splendid Feather Moss Grimmia pulvinata Grey-cushioned Grimmia Hypnum circinale Coiled-Leaf Moss Leucolepis acanthoneuron Menzie’s Tree Moss, Umbrella Moss, Palm-Tree Moss Plagiomnium insigne Badge Moss, Coastal Leafy Moss Pseudotaxiphyllum elegans Small-Flat Moss Rhizomnium glabrescens Fan Moss Rhytidiadelphus loreus Lanky Moss, Loreus Goose Neck Moss Rhytidiadelphus squarrosum Springy Turf-Moss, Square Goose Neck Moss Rhytidiadelphus triquetrus Electrified Cat-Tail Moss, Goose Necked Moss Rhytidiopsus robusta Robust mountain moss Schistostega pennata Goblin’s Gold, Luminous Moss Polytrichopsida: Atrichum Atrichum Moss , Crane’s Bill Moss (for Atrichum selwynii) Pogonatum contortum Contorted Pogonatum Moss Polytrichum commune Common Hair Cap Moss Polytrichum piliferum Bristly Haircap Moss Andreaeopsida Andreaea nivalis Granite moss, Lantern moss, Snow Rock Moss Sphagnopsida: Sphagnum capillifolium Red Bog Moss, Small Red Peat Moss Sphagnum papillosum Fat Bog Moss, Papillose sphagnum Sphagnum squarrosum Shaggy Sphagnum, Spread- Leaved Peat Moss Takakiopsida: Takakia lepidoziooides Impossible -

Coptis Trifolia Conservation Assessment
CONSERVATION ASSESSMENT for Coptis trifolia (L.) Salisb. Originally issued as Management Recommendations December 1998 Marty Stein Reconfigured-January 2005 Tracy L. Fuentes USDA Forest Service Region 6 and USDI Bureau of Land Management, Oregon and Washington CONSERVATION ASSESSMENT FOR COPTIS TRIFOLIA Table of Contents Page List of Tables ................................................................................................................................. 2 List of Figures ................................................................................................................................ 2 Summary........................................................................................................................................ 4 I. NATURAL HISTORY............................................................................................................. 6 A. Taxonomy and Nomenclature.......................................................................................... 6 B. Species Description ........................................................................................................... 6 1. Morphology ................................................................................................................... 6 2. Reproductive Biology.................................................................................................... 7 3. Ecological Roles ............................................................................................................. 7 C. Range and Sites -

Riparian Bryophytes of the H. J. Andrews Experimental Forest in the Western Cascades, Oregon
The Bryologist 99(2), pp. 226-235 Copyright © 1996 by the American Bryological and Lichenological Society, Inc. Riparian Bryophytes of the H. J. Andrews Experimental Forest in the Western Cascades, Oregon BENGT GUNNAR JONSSON I Department of Forest Science, Oregon State University, Corvallis, OR 97331 Abstract. The knowledge of the distribution and habitat demands for bryophytes in the Pacific Northwest is scarce, and few published quantitative accounts of the flora are present. The present paper includes habitat description, elevational range, substrate preference, and frequency esti- mates for more than 130 riparian mosses and liverworts found in old-growth Pseudotsuga-Tsuga forests of the H. J. Andrews Experimental Forest, Oregon. The data are based on 360 samples distributed among 42 sites covering 1st to 5th order streams and 420 to 1250 m. TWINSPAN analysis resulted in 6 sample groups, representing samples from different elevations, geomorphic surfaces, and stream sizes. The most common mosses are Eurhynchium oreganum, Isothecium stoloniferum, Hypnum circinale, and Dicranum fuscescens. Among the hepatics Scapania bolan- deri, Cephalozia lunulifolia, and Porella navicularis are the most abundant species. Most species are rare at both site and sampte gtot (ever; and this is especially true for acrocarps where more than one-third of the observed species occurred in only one or two sites orland samples. Four of the occurring species (i.e., Antitrichia curtipendula, Buxbaumia piperi, Douinia ovata, and Ptili- dium californicum) are listed for special management and/or regional surveys. Bryophytes constitute an important and conspic- a wide range of different substrates, disturbance uous component of old-growth forests in the Pacific patterns, and a moist microclimate are important Northwest (Lesica et al. -

Flora of New Zealand Mosses
FLORA OF NEW ZEALAND MOSSES BRACHYTHECIACEAE A.J. FIFE Fascicle 46 – JUNE 2020 © Landcare Research New Zealand Limited 2020. Unless indicated otherwise for specific items, this copyright work is licensed under the Creative Commons Attribution 4.0 International licence Attribution if redistributing to the public without adaptation: "Source: Manaaki Whenua – Landcare Research" Attribution if making an adaptation or derivative work: "Sourced from Manaaki Whenua – Landcare Research" See Image Information for copyright and licence details for images. CATALOGUING IN PUBLICATION Fife, Allan J. (Allan James), 1951- Flora of New Zealand : mosses. Fascicle 46, Brachytheciaceae / Allan J. Fife. -- Lincoln, N.Z. : Manaaki Whenua Press, 2020. 1 online resource ISBN 978-0-947525-65-1 (pdf) ISBN 978-0-478-34747-0 (set) 1. Mosses -- New Zealand -- Identification. I. Title. II. Manaaki Whenua-Landcare Research New Zealand Ltd. UDC 582.345.16(931) DC 588.20993 DOI: 10.7931/w15y-gz43 This work should be cited as: Fife, A.J. 2020: Brachytheciaceae. In: Smissen, R.; Wilton, A.D. Flora of New Zealand – Mosses. Fascicle 46. Manaaki Whenua Press, Lincoln. http://dx.doi.org/10.7931/w15y-gz43 Date submitted: 9 May 2019 ; Date accepted: 15 Aug 2019 Cover image: Eurhynchium asperipes, habit with capsule, moist. Drawn by Rebecca Wagstaff from A.J. Fife 6828, CHR 449024. Contents Introduction..............................................................................................................................................1 Typification...............................................................................................................................................1 -

Volume 1, Chapter 2-7: Bryophyta
Glime, J. M. 2017. Bryophyta – Bryopsida. Chapt. 2-7. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological Ecology. Ebook 2-7-1 sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 10 January 2019 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 2-7 BRYOPHYTA – BRYOPSIDA TABLE OF CONTENTS Bryopsida Definition........................................................................................................................................... 2-7-2 Chromosome Numbers........................................................................................................................................ 2-7-3 Spore Production and Protonemata ..................................................................................................................... 2-7-3 Gametophyte Buds.............................................................................................................................................. 2-7-4 Gametophores ..................................................................................................................................................... 2-7-4 Location of Sex Organs....................................................................................................................................... 2-7-6 Sperm Dispersal .................................................................................................................................................. 2-7-7 Release of Sperm from the Antheridium..................................................................................................... -

Bryophytes from the Cape Verde Islands
123 Tropical Bryology 12: 123-153, 1996 Bryophytes from the Cape Verde Islands Jan-Peter Frahm1, Anja Lindlar1, Philip Sollman2, Eberhard Fischer1 1 Botanisches Institut der Rheinischen Friedrich-Wilhelms-Universität, Meckenheimer Allee 170, 53115 Bonn, Germany 2 Von Weberstraat 32, 6904 KD Zevenaar, The Netherlands Abstract: Almost 450 specimens of bryophytes, so far the largest collection of bryophytes ever made on the Cape Verde Islands, were collected in 1995 by the second author on the major islands of the archipelago. Twenty seven species (3 hepatics, 24 mosses) are reported as new to the Cape Verde Islands: Lejeunea ulicina (Tayl.) Gottsche et al., Riccia cavernosa Hoffm. emend. Raddi, Targionia hypophylla L., Barbula cf. consanguinea (Thwait. & Mitt.) Jaeg., Barbula unguiculata Hedw., Brachymenium exile (Dozy & Molk.) Bosch. & Lac., Bryoerythrophyllum ferruginascens (Stirt.) Giac., Bryoerythrophyllum inaequalifolium (Tayl.) Zander, Bryum cellulare Hook., Chenia leptophylla (C. Müll.) Zander, Desmatodon bogosicus C. Müll., Didymodon australasiae (Hook. & Grev.) Zander, Didymodon maschalogena (Ren. & Card.) Broth. (Didymodon michiganensis [Steere] K. Saito), Didymodon vinealis (Brid.) Zander var. flaccidus (B.S.G.) Zander, Eurhynchium meridionale (B.S.G.) De Not., Eurhynchium speciosum (Brid.) Jur., Fissidens sciophyllus Mitt., F. bogosicus C. Müll., F. flaccidus Mitt., F. helictocaulos C. Müll., Gymnostomiella cf. vernicosa (Hook.) Fleisch., Gymnostomum calcareum Nees & Hornsch., Hyophila involuta (Hook.) Jaeg., Orthotrichum diaphanum Brid., Tortula cuneifolia (With.) Turn., Tortula laevipila (Brid.) Schwaegr. and Weissia microstoma (Hedw.) C. Müll. The doubtful record of Marchantia paleacea Bertol. could be confirmed. Numerous species are recorded as new to single islands. Tortula pierrotii Biz. described from Tanzania has proved to be synyomous with Bryoerythrophyllum inaequalifolium. Didymodon maschalogena (Ren. -

The Biflavonoid Pattern of Rhytidiadelphus Squarrosus (Hedw.) Warnst
The Biflavonoid Pattern of Rhytidiadelphus squarrosus (Hedw.) Warnst. Tassilo Seeger, Hans Dietmar Zinsmeister FB 13, Botanik, Universität des Saarlandes, D-6600 Saarbrücken, Bundesrepublik Deutschland Hans Geiger Institut für Chemie der Universität Hohenheim, D-7000 Stuttgart 70, Bundesrepublik Deutschland Z. Naturforsch. 45c, 583-586 (1990); received January 22, 1990 Dedicated to Dr. Ella Campbell, Massey University, Palmerston North, New Zealand, on the occasion of her 80th birthday Mosses, Hylocomiaceae, Rhytidiadelphus squarrosus, Biflavonoids, 5'-Hydroxyrobustaflavone From Rhytidiadelphus squarrosus the hitherto unknown biflavone 5'-hydroxyrobusta- flavone and the three known biflavonoids 5'-hydroxyamentoflavone,5',3"'-dihydroxyamento- flavone and 2,3-dihydro-5'-hydroxyamentoflavone have been isolated. Introduction 0H Various biflavonoids have been isolated so far from eleven species of seven different moss fami lies, namely Dicranaceae [1-3], Grimmiaceae [4], Bryaceae [5], Mniaceae [4,6], Bartramiaceae [7], Leucodontaceae [4] and Hylocomiaceae [4,8 ]. The most common moss biflavonoids are di mers of luteolin and their 2,3- or 2",3"-dihydro-de- rivatives. Biflavonoids containing monomers oth 5'-Hydroxyamentoflavone (1) R = H 5'-3"'-Dihydroxyamentoflavone (2) R = OH er than luteolin or eriodictyol (= 2,3-dihydroluteo- lin) have been found until now only in species of Bryaceae and Mniaceae [9], both belonging to the 0H suborder Bryineae Fleisch. [10]. Here the isolation of three biflavonoids, containing an apigenin moiety, from Rhytidiadelphus squarrosus which belongs to the Hypnineae Fleisch. [10] is reported. Results and Discussion From Rhytidiadelphus squarrosus four biflavon 5'-Hydroxyrobustaflavone (3) R = H oids 1, 2, 3 and 4 were isolated. 1, 2 and 4 are 5',3"'-Dihydroxyrobustaflavone (5) R = OH known compounds. -

Rhytidiadelphus Squarrosus
Rhytidiadelphus squarrosus Britain 1990–2013 2416 1950–1989 286 pre-1950 5 Ireland 1990–2013 590 1950–1989 180 pre-1950 4 robust moss with a wide ecological tolerance, occurring by heavy grazing and mowing, but is invariably absent from Aon all but the most acid soils in a variety of grassy reseeded and agriculturally improved fields. Altitudinal habitats, including sheep pastures, roadside and trackside range: 0–1170 m. verges, light woodland and scrub, dunes, streamsides, ditches and marshes. It also occurs among ericaceous shrubs Dioicous; capsules are rare in the lowlands but more and in flushed turf on heath and moorland. It is a very frequent in the north and west, mature in winter and early common species of lawns (where it attracts the attention and spring. Its ubiquity is remarkable, considering the rarity of often the disapproval of gardeners!), as well as churchyards, capsules and lack of gemmae. Presumably it is dispersed parkland and other places that are regularly mown and vegetatively by stem and leaf fragments. not heavily fertilised, and likewise it occurs in low-lying unimproved pastures. Regular associates include Lophocolea Plants from woodland often have a lax habit resembling bidentata, Brachythecium rutabulum, Calliergonella cuspidata, R. subpinnatus and the separation of the two species is Kindbergia praelonga and Pseudoscleropodium purum. It sometimes difficult. It is possible that there may be a few ascends to high altitudes in mountain grassland and Nardus recording errors in woodland habitats, but with little impact snowbeds, where associates include Hylocomium splendens, on the overall accuracy of the map. Pleurozium schreberi and Rhytidiadelphus loreus. -

Bryophyte Diversity and Vascular Plants
DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 75 BRYOPHYTE DIVERSITY AND VASCULAR PLANTS NELE INGERPUU TARTU 2002 DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 75 DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 75 BRYOPHYTE DIVERSITY AND VASCULAR PLANTS NELE INGERPUU TARTU UNIVERSITY PRESS Chair of Plant Ecology, Department of Botany and Ecology, University of Tartu, Estonia The dissertation is accepted for the commencement of the degree of Doctor philosophiae in plant ecology at the University of Tartu on June 3, 2002 by the Council of the Faculty of Biology and Geography of the University of Tartu Opponent: Ph.D. H. J. During, Department of Plant Ecology, the University of Utrecht, Utrecht, The Netherlands Commencement: Room No 218, Lai 40, Tartu on August 26, 2002 © Nele Ingerpuu, 2002 Tartu Ülikooli Kirjastuse trükikoda Tiigi 78, Tartu 50410 Tellimus nr. 495 CONTENTS LIST OF PAPERS 6 INTRODUCTION 7 MATERIAL AND METHODS 9 Study areas and field data 9 Analyses 10 RESULTS 13 Correlation between bryophyte and vascular plant species richness and cover in different plant communities (I, II, V) 13 Environmental factors influencing the moss and field layer (II, III) 15 Effect of vascular plant cover on the growth of bryophytes in a pot experiment (IV) 17 The distribution of grassland bryophytes and vascular plants into different rarity forms (V) 19 Results connected with nature conservation (I, II, V) 20 DISCUSSION 21 CONCLUSIONS 24 SUMMARY IN ESTONIAN. Sammaltaimede mitmekesisus ja seosed soontaimedega. Kokkuvõte 25 < TÄNUSÕNAD. Acknowledgements 28 REFERENCES 29 PAPERS 33 2 5 LIST OF PAPERS The present thesis is based on the following papers which are referred to in the text by the Roman numerals. -

Biodiversity, Conservation and Cultural History
Sycamore maple wooded pastures in the Northern Alps: Biodiversity, conservation and cultural history Inauguraldissertation der Philosophisch-naturwissenschaftlichen Fakultät der Universität Bern vorgelegt von Thomas Kiebacher von Brixen (Italien) Leiter der Arbeit: Prof. Dr. Christoph Scheidegger Dr. Ariel Bergamini PD Dr. Matthias Bürgi WSL Swiss Federal Research Institute, Birmensdorf Sycamore maple wooded pastures in the Northern Alps: Biodiversity, conservation and cultural history Inauguraldissertation der Philosophisch-naturwissenschaftlichen Fakultät der Universität Bern vorgelegt von Thomas Kiebacher von Brixen (Italien) Leiter der Arbeit: Prof. Dr. Christoph Scheidegger Dr. Ariel Bergamini PD Dr. Matthias Bürgi WSL Swiss Federal Research Institute, Birmensdorf Von der Philosophisch-naturwissenschaftlichen Fakultät angenommen. Bern, 13. September 2016 Der Dekan: Prof. Dr. Gilberto Colangelo Meinen Eltern, Frieda und Rudolf Contents Abstract ................................................................................................................................................... 9 Introduction ........................................................................................................................................... 11 Context and aims ............................................................................................................................... 13 The study system: Sycamore maple wooded pastures ..................................................................... 13 Biodiversity ....................................................................................................................................... -

Liverworts Mosses
LIVERWORTS LIVERWORTS MOSSES Heller’s Notchwort (Anastrophyllum hellerianum) Hatcher’s Paw-wort (Barbilophozia hatcheri) Key features for identifying Key features for identifying liverworts Mosses Growth form. There are two sorts of liverworts; leafy Growth form. Leaving aside the very distinctive bog- liverworts have a stem and leaves and resemble a mosses (Sphagnum), mosses can be split into two moss, whereas thallose or thalloid liverworts have a groups, acrocarpous and pleurocarpous. There is a simple strap of tissue with no stem or leaves. Leafy technical difference between these two forms but in liverworts can form erect cushions and turfs while some practical terms, acrocarps usually have erect stems are creeping and closely apressed to rock or tree. The and grow in cushions or turfs while pleurocarps tend size of the plant is also important; a number of oceanic Michael Lüth to grow with main stems parallel to the ground (or rock Leafy liverwort NS; size: very small and forming thin patches Leafy liverwort; size: small to medium-sized forming tight liverworts are very, very small. Above - Left: acrocarpous Scott’s Fork moss (Dicranum Above - Left: a thallose liverwort, Overleaf Pellia (Pellia or tree trunk) and form wefts. The often dense growth of upright stems; colour: yellow or yellow green with shoot patches of erect stems; colour: mid or yellow green, often with Leaf shape. This is all-important in leafy liverworts and scottianum) ; right: pleurocarpous Larger Mouse-tail epiphylla) with fruits; right: a leafy liverwort with round form of acrocarps means that their sparse branches tips red with gemmae; leaves: tiny with two lobes but hardly some shoot tips red with gemmae; leaves: rounded-rectangular leaves, Autumn Flapwort (Jamesoniella autumnalis). -

(Limpr.) Warnst
ISSN 2336-3193 Acta Mus. Siles. Sci. Natur., 68: 123-134, 2019 DOI: 10.2478/cszma-2019-0012 Published: online 1 July 2019, print July 2019 On moss genera Hylocomiadelphus Ochyra & Stebel and Rhytidiadelphus (Limpr.) Warnst. Michael S. Ignatov, Elena A. Ignatova, Oxana I. Kuznetsova & Vladimir E. Fedosov On moss genera Hylocomiadelphus Ochyra & Stebel and Rhytidiadelphus (Limpr.) Warnst. – Acta Mus. Siles. Sci. Natur., 68: 123-134, 2019. Abstract: The moss family Hylocomiaceae is studied for the generic level taxonomy within a molecular phylogenetic approach. We confirm segregating of the species formerly known as Rhytidiadelphus triquetrus to Hylocomiadelphus Ochyra & Stebel from the genus Rhytidiadelphus . Hylocomiadelphus forms a clade with Loeskeobryum and Meteoriella , sharing with both genera cordate to auriculate leaf bases and non-squarrose leaves, and with the former undulate leaves and reticulate exostome teeth ornamentation. However, Loeskeobryum differs from Hylocomiadelphus in having paraphyllia, while Meteoriella has a straight capsule and reduced peristome, likely caused by its epiphytic ecology. In the group of species closely related to R. squarrosus , in addition to R. subpinnatus , the third species with North Pacific distribution is described as R. pacificum . Macrothamnium is found deeply nested in Rhytidiadelphus , although no nomenclatural implications are suggested at the moment due to insufficient sampling. Hylocomium splendens var. splendens and var. obtusifolium were found intermingled in the phylogenetic tree, indicating no correlation between morphology and variation in ITS region, thus supporting a view that these taxa are merely environmentally induced morphs. Key words: bryophytes, Rhytidiadelphus , Hylocomiadelphus , Hylocomium , Hylocomiaceae, taxono- my, molecular phylogeny, phytogeography, new species, new genera, nomenclature Introduction Despite that the moss family Hylocomiaceae M.