Amphibolurus Muricatus):Anagamidlizard with Temperature-Dependent Sex Determination
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

This Article Appeared in a Journal Published by Elsevier. the Attached Copy Is Furnished to the Author for Internal Non-Commerci
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights Author's personal copy Molecular Phylogenetics and Evolution 71 (2014) 149–156 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev A multi-locus molecular phylogeny for Australia’s iconic Jacky Dragon (Agamidae: Amphibolurus muricatus): Phylogeographic structure along the Great Dividing Range of south-eastern Australia ⇑ Mitzy Pepper a, , Marco D. Barquero b, Martin J. Whiting b, J. Scott Keogh a a Division of Evolution, Ecology and Genetics, Research School of Biology, The Australian National University, Canberra, Australia b Department of Biological Sciences, Macquarie University, Sydney, Australia article info abstract Article history: Jacky dragons (Amphibolurus muricatus) are ubiquitous in south-eastern Australia and were one of the Received 25 June 2013 -

An Annotated Type Catalogue of the Dragon Lizards (Reptilia: Squamata: Agamidae) in the Collection of the Western Australian Museum Ryan J
RECORDS OF THE WESTERN AUSTRALIAN MUSEUM 34 115–132 (2019) DOI: 10.18195/issn.0312-3162.34(2).2019.115-132 An annotated type catalogue of the dragon lizards (Reptilia: Squamata: Agamidae) in the collection of the Western Australian Museum Ryan J. Ellis Department of Terrestrial Zoology, Western Australian Museum, Locked Bag 49, Welshpool DC, Western Australia 6986, Australia. Biologic Environmental Survey, 24–26 Wickham St, East Perth, Western Australia 6004, Australia. Email: [email protected] ABSTRACT – The Western Australian Museum holds a vast collection of specimens representing a large portion of the 106 currently recognised taxa of dragon lizards (family Agamidae) known to occur across Australia. While the museum’s collection is dominated by Western Australian species, it also contains a selection of specimens from localities in other Australian states and a small selection from outside of Australia. Currently the museum’s collection contains 18,914 agamid specimens representing 89 of the 106 currently recognised taxa from across Australia and 27 from outside of Australia. This includes 824 type specimens representing 45 currently recognised taxa and three synonymised taxa, comprising 43 holotypes, three syntypes and 779 paratypes. Of the paratypes, a total of 43 specimens have been gifted to other collections, disposed or could not be located and are considered lost. An annotated catalogue is provided for all agamid type material currently and previously maintained in the herpetological collection of the Western Australian Museum. KEYWORDS: type specimens, holotype, syntype, paratype, dragon lizard, nomenclature. INTRODUCTION Australia was named by John Edward Gray in 1825, The Agamidae, commonly referred to as dragon Clamydosaurus kingii Gray, 1825 [now Chlamydosaurus lizards, comprises over 480 taxa worldwide, occurring kingii (Gray, 1825)]. -

Reproductive Ecology of the Mountain Dragon, Rankin/A (Tympanocryptis) Diemensis (Reptilia: Squamata: Agamidae) in Tasmania
Papers and Proceedings ofthe Royal Society of Tasmania, Volume 139, 2005 23 REPRODUCTIVE ECOLOGY OF THE MOUNTAIN DRAGON, RANKIN/A (TYMPANOCRYPTIS) DIEMENSIS (REPTILIA: SQUAMATA: AGAMIDAE) IN TASMANIA by Jemina Stuart-Smith, Roy Swain and Andrew Welling (with three tables and two text-figures) Stuart-Smith, J., Swain, R. & Welling, A. 2005 (16:xii): Reproductive ecology of the Mountain Dragon, Rankinia (Tympanocryptis) diemensis (Reptilia: Squamata: Agamidae) in Tasmania. Papers and Proceedings of the Royal Society of Tasmania 139: 23-28. https://doi.org/10.26749/rstpp.139.23 ISSN 0080-4703. School of Zoology, University of Tasmania, Private Bag 5, Tasmania, 7001 Australia.(J.S-S.*, R.S., AW). *Author for correspondence. The mountain dragon, Rankinia (Tympanocryptis) diemensis (Gray, 1841), is the only member of the Agamidae in Tasmania. It occurs in some of the coldest regions occupied by any dragon in Australia, and is found in a variety of habitats ranging from coastal heath to alpine scrub. This paper examines the reproductive ecology of R. diemensis in the most southerly range of its distribution, providing baseline data on timing of reproductive events, reproductive cycles, nesting behaviour and ovipositioning, clutch characteristics and incubation conditions. Winter torpor lasts approximately seven months with males emerging in early September and spermatogenesis occurring from September-November. Females emerge later, with vitcllogenesis occurring from September-December. Gravid females may be found between October and January, but females are non-vitcllogenic from late December until the following season. The firstclutch is typically laid from October-December, with a variable clutch size (2-11 eggs). Females store sperm and a second clutch may be laid fiveweeks after the first. -

KENNETH A. NAGY PUBLICATIONS up to JUNE 2016
KENNETH A. NAGY PUBLICATIONS Up to JUNE 2016 For Open Access pdf, click link. For copyrighted reprint pdfs, please email your request to Ken Nagy at [email protected]. I will be happy to share a copy of the article with individual colleagues, when permissible under copyright law. 2016 Nagy, K.A., G. Kuchling, L.S. Hillard, and B.T. Henen. (2016) Weather and sex ratios of head-started Agassiz’s desert tortoise Gopherus agassizii juveniles hatched in natural habitat enclosures. Endangered Species Research 30:145- 155. (Research article) (Link to pdf) Ellsworth, E., M.R. Boudreau, K. Nagy, J.L. Rachlow, and D.L. Murray. (2016). Differential sex-related winter energetics in free-ranging snowshoe hAres (Lepus americanus). Canadian Journal of Zoology 94:115-121. (Research article) (link to pdf) 2015 Nagy, K.A., S.Hillard, S. Dickson, and D.J. Morafka. (2015). Effects of artificial rain on survivorship, body condition, and growth of head-started desert tortoises (Gopherus agassizii) released to the open desert. Herpetological Conservation and Biology 10:535-549. (Research article) (Link to pdf) Nagy, K.A., L.S. Hillard, M.W. Tuma, and D.J. Morafka. (2015). Head-started desert tortoises (Gopherus agassizii): Movements, survivorship and mortality causes following their release. Herpetological Conservation and Biology 10:203-215. (Research article) (link to pdf) 2014 Gienger, C.M., C.R. Tracy, and K.A. Nagy. (2014). Life in the slow lane: GilA Monsters have low rates of energy use and water flux. Copeia 2014:279-287. (Research article) (email Nagy for pdf) 2012 Nagy, K.A., and G.G. -

Testing the Relevance of Binary, Mosaic and Continuous Landscape Conceptualisations to Reptiles in Regenerating Dryland Landscapes
Testing the relevance of binary, mosaic and continuous landscape conceptualisations to reptiles in regenerating dryland landscapes Melissa J. Bruton1, Martine Maron1,2, Noam Levin1,3, Clive A. McAlpine1,2 1The University of Queensland, Landscape Ecology and Conservation Group, School of Geography, Planning and Environmental Management, St Lucia, Australia 4067 2The University of Queensland, ARC Centre of Excellence for Environmental Decisions, St. Lucia, Australia 4067 3Hebrew University of Jerusalem, Department of Geography, Mt. Scopus, Jerusalem, Israel, 91905 Corresponding author: [email protected] Ph: (+61) 409 875 780 The final publication is available at Springer via http://dx.doi.org/10.1007/s10980-015-0157-9 Abstract: Context: Fauna distributions are assessed using discrete (binary and mosaic) or continuous conceptualisations of the landscape. The value of the information derived from these analyses depends on the relevance of the landscape representation (or model) used to the landscape and fauna of interest. Discrete representations dominate analyses of landscape context in disturbed and regenerating landscapes; however within-patch variation suggests that continuous representations may help explain the distribution of fauna in such landscapes. Objectives: We tested the relevance of binary, mosaic, and continuous conceptualisations of landscape context to reptiles in regenerating dryland landscapes. Methods: For each of thirteen reptile groups, we compared the fit of models consisting of one landscape composition and one landscape heterogeneity variable for each of six landscape representations (2 x binary, 2 x mosaic, and 2 x continuous), at three buffer distances. We used Akaike weights to assess the relative support for each model. Maps were created from Landsat satellite images. -

Australian Capital Territory & Yass Valley Reptiles
Australian Capital Territory & Yass Valley Reptiles Geoff Robertson 4 June 2017 Turtle (1-2 species) Lizard Gecko (2) Legless lizard (4-5) Dragon (6) Gonna (2) Skink (32-35) Egernia group - chunky (9) Eugongylus group - striped (9-10) Sphenomorphus group - red (14) Snakes Blind (1) Total species = 57-66 Python (1?) Elapid (9-12) Murray (short-neck) turtle (above left) Emydura macquarii Eastern long-neck turtle (above right) Chelodina longicollis Lizards Monitors (goannas) Dragons Skinks Geckos Measuring length lizards & snakes: Legless TL - total length. lizards SVL - tip of the nose (snout) to vent. Dragons Physignathus lesueurii, Australia: 11 genera, 70 species. SVL 245 mm ACT/Yass: 5 genera, 6 species Common: Pogona barbata, Water dragon (top), Aust: 1 SVL 250 mm species. Eastern bearded dragon (bottom), Aust: 6 species. Jacky lizard or tree dragon. Three similar dragons Jacky (top), Nobbi (middle) and Mountain (bottom). SVL is Jacky - 120mm Nobbi - 84mm Mountain - 82mm However, all dragons (including water & bearded ) look similar when young. Photos top to bottom - Amphibolurus mutinous and A. nobbi, & Rankina diemensis. by Warren Saunders, John Wombey and Roger Farrow. Grassland earless dragon Eight earless dragon species in Australia. GED found in some natural grassland sites, climbs tussock grasses otherwise a ground dweller. Unlike other dragons, Tail slight/curls. Populations - ACT & southern Monaro. GED thought extinct, re-discovered, crashed in drought, captive breeding, future? Tympanocryptis pinguicolla. SVL 55mm. Bottom photo: Margaret Ning. Jacky dragon Bearded dragon Water dragon Nobbi dragon Mountain dragon Grassland earless dragon Gecko Australia: 18 genera & 111 species ACT/Yass: 2 species Stone gecko (right) Marble gecko (below) Diplodactylus vittatus, SVL 50mm Photo John Wombey Christinus marmaratus, SVL 70mm Photo Margaret Ning Legless lizards Related to geckos Australia: 7 genera & 38 species ACT/Yass: 2 genera & 5 species Unlike snakes: Lialis burtonis, SVL170mm Photo John Wombey • Large fleshy tongue (cf forked). -

Fauna of Australia 2A
FAUNA of AUSTRALIA 26. BIOGEOGRAPHY AND PHYLOGENY OF THE SQUAMATA Mark N. Hutchinson & Stephen C. Donnellan 26. BIOGEOGRAPHY AND PHYLOGENY OF THE SQUAMATA This review summarises the current hypotheses of the origin, antiquity and history of the order Squamata, the dominant living reptile group which comprises the lizards, snakes and worm-lizards. The primary concern here is with the broad relationships and origins of the major taxa rather than with local distributional or phylogenetic patterns within Australia. In our review of the phylogenetic hypotheses, where possible we refer principally to data sets that have been analysed by cladistic methods. Analyses based on anatomical morphological data sets are integrated with the results of karyotypic and biochemical data sets. A persistent theme of this chapter is that for most families there are few cladistically analysed morphological data, and karyotypic or biochemical data sets are limited or unavailable. Biogeographic study, especially historical biogeography, cannot proceed unless both phylogenetic data are available for the taxa and geological data are available for the physical environment. Again, the reader will find that geological data are very uncertain regarding the degree and timing of the isolation of the Australian continent from Asia and Antarctica. In most cases, therefore, conclusions should be regarded very cautiously. The number of squamate families in Australia is low. Five of approximately fifteen lizard families and five or six of eleven snake families occur in the region; amphisbaenians are absent. Opinions vary concerning the actual number of families recognised in the Australian fauna, depending on whether the Pygopodidae are regarded as distinct from the Gekkonidae, and whether sea snakes, Hydrophiidae and Laticaudidae, are recognised as separate from the Elapidae. -

Nelson Et Al No Highlight-1 Edit Changes
1 Receiver psychology and the design of the deceptive caudal luring signal of the death 2 adder 3 Ximena J. Nelsona,b* Daniel T. Garnettc, 1 Christopher S. Evansb, 2 4 5 aSchool of Biological Sciences, University of Canterbury 6 bCentre for the Integrative Study of Animal Behaviour, Macquarie University 7 cDepartment of Biological Sciences, Macquarie University 8 Received 6 July 2009 9 Initial acceptance 4 September 2009 10 Final acceptance 30 November 2009 11 MS. number: 09-00453R 12 13 *Correspondence: X. J. Nelson, School of Biological Sciences, University of Canterbury, 14 Private Bag 4800, Christchurch, New Zealand. 15 E-mail address: [email protected] (X. J. Nelson). 16 1D. T. Garnett is at the Department of Biological Sciences, Macquarie University, Sydney, 17 NSW 2109 Australia. 18 2C. S. Evans is at the Centre for the Integrative Study of Animal Behaviour, Macquarie 19 University, Sydney, NSW 2109 Australia. 20 1 21 Signal design can reflect the sensory properties of receivers. The death adder, Acanthophis 22 antarcticus, attracts prey by wriggling the distal portion of its tail (caudal luring). To 23 understand the design of this deceptive signal, we explored perceptual processes in a 24 representative prey species: the Jacky dragon, Amphibolurus muricatus. We used 3D 25 animations of fast and slow death adder luring movements against different backgrounds, to 26 test the hypothesis that caudal luring mimics salient aspects of invertebrate prey. Moving 27 stimuli elicited predatory responses, especially against a conspicuous background. To identify 28 putative models for caudal luring, we used an optic flow algorithm to extract velocity values 29 from video sequences of 61 moving invertebrates caught in lizard territories, and compared 30 these to the velocity values of death adder movements. -

Wollongong Campus Fauna Species List
FACTSHEET Wollongong Campus Fauna Species List The Wollongong Campus landscape provides habitat for a diverse range of animals. Animals that are known or potentially occur on campus are listed below. Key: # Introduced species, * Listed as a Vulnerable or threatened species in NSW, C- common, E - exotic, Res- resident, Br - breeds at UOW, M - migratory, V – visitor, P – potentially occurs onsite AMPHIBIANS Bleating tree frog Litoria dentate Res Eastern dwarf tree frog Litoria fallax Res Eastern froglet Crinia signifera C, Res Leaf green tree frog Litoria nudidigita Res Peron’s tree frog Litoria peronei Res Striped marsh frog Limnodynastes peronei Res Tyler’s tree frog Litoria tyleri Res Verreaux’s tree frog Litoria verreauxi Res REPTILES Common garden skink Lampropholis guichenoti C, Res Diamond python Morelia spilota Res Eastern blue tongue lizard Tilliqua scincoides C, Res Eastern long - necked turtle Chelondina longicollis Res Eastern small - eyed snake Cryptophis nigrescens Res Eastern tiger snake Notechis scutatus Res (escarpment) Eastern water dragon Physignathus lesurerii C, Res Eastern water skink Eulamprus quoyii C, Res Jacky dragon Amphibolurus muricatus Res Golden crowned snake Cacophis squamulosus Res Macquarie turtle Emydura macquarii Res Marsh (swamp) snake Hemiaspis signata Res Mustard bellied snake Drysdalia rhodogaster V Red - bellied black snake Pseudechis porphriacus Res Sheoak skink Cyclodomorphus michaeli Res Three - toed skink Saiphos equalis Res Weasel skink Saproscincus mustelinus Res MAMMALS Bush rat Rattus fuscipes -
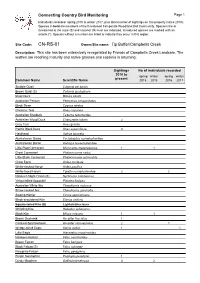
CN-RS-01 Owner/Site Name: Tip Buffer/Campbells Creek
Connecting Country Bird Monitoring Page 1 Individuals recorded spring 2015 to winter 2017, plus total number of sightings on this property (since 2010). Species in bold are members of the threatened Temperate Woodland Bird Community. Species rare or threatened at the state (S) and national (N) level are indicated. Introduced species are marked with an asterix (*). Species without a number are listed to indicate they occur in this region. Site Code: CN-RS-01 Owner/Site name: Tip Buffer/Campbells Creek Description: This site has been extensively revegetated by Friends of Campbells Creek Landcare. The wattles are reaching maturity and native grasses and cassinia is returning. Sightings No of individuals recorded 2010 to spring winter spring winter present Common Name Scientific Name 2015 2016 2016 2017 Stubble Quail Coturnix pectoralis Brown Quail (S) Coturnix ypsilophora Musk Duck Biziura lobata Australian Pelican Pelecanus conspicillatus Black Swan Cygnus atratus Chestnut Teal Anas castanea Australian Shelduck Tadorna tadornoides Australian Wood Duck Chenonetta jubata 2 Grey Teal Anas gracilis Pacific Black Duck Anas superciliosa 3 Hardhead Aythya australis Australasian Grebe Tachybaptus novaehollandiae Australasian Darter Anhinga novaehollandiae Little Pied Cormorant Microcarbo melanoleucos 1 1 Great Cormorant Phalacrocorax carbo Little Black Cormorant Phalacrocorax sulcirostris Great Egret Ardea modesta White-necked Heron Ardea pacifica White-faced Heron Egretta novaehollandiae 2 2 Nankeen Night-Heron (S) Nycticorax caledonicus Yellow-billed -

A Molecular Phylogenetic Study of Ecological Diversification in the Australian Lizard Genus Ctenophorus
JEZ Mde 2035 JOURNAL OF EXPERIMENTAL ZOOLOGY (MOL DEV EVOL) 291:339–353 (2001) A Molecular Phylogenetic Study of Ecological Diversification in the Australian Lizard Genus Ctenophorus JANE MELVILLE,* JAMES A. SCHULTE II, AND ALLAN LARSON Department of Biology, Washington University, St. Louis, Missouri 63130 ABSTRACT We present phylogenetic analyses of the lizard genus Ctenophorus using 1,639 aligned positions of mitochondrial DNA sequences containing 799 parsimony-informative charac- ters for samples of 22 species of Ctenophorus and 12 additional Australian agamid genera. Se- quences from three protein-coding genes (ND1, ND2, and COI) and eight intervening tRNA genes are examined using both parsimony and maximum-likelihood analyses. Species of Ctenophorus form a monophyletic group with Rankinia adelaidensis, which we suggest placing in Ctenophorus. Ecological differentiation among species of Ctenophorus is most evident in the kinds of habitats used for shelter. Phylogenetic analyses suggest that the ancestral condition is to use burrows for shelter, and that habits of sheltering in rocks and shrubs/hummock grasses represent separately derived conditions. Ctenophorus appears to have undergone extensive cladogenesis approximately 10–12 million years ago, with all three major ecological modes being established at that time. J. Exp. Zool. (Mol. Dev. Evol.) 291:339–353, 2001. © 2001 Wiley-Liss, Inc. The agamid lizard genus Ctenophorus provides ecological categories based on whether species abundant opportunity for a molecular phylogenetic shelter in rocks, burrows, or vegetation. Eight spe- study of speciation and ecological diversification. cies of Ctenophorus are associated with rocks: C. Agamid lizards show a marked radiation in Aus- caudicinctus, C. decresii, C. fionni, C. -

Technique for Measuring Speed and Visual Motion Sensitivity in Lizards
Psicológica (2008), 29, 133-151. Technique for measuring speed and visual motion sensitivity in lizards Kevin L. Woo* and Darren Burke Macquarie University (Australia) Testing sensory characteristics on herpetological species has been difficult due to a range of properties related to physiology, responsiveness, performance ability, and the type of reinforcer used. Using the Jacky lizard as a model, we outline a successfully established procedure in which to test the visual sensitivity to motion characteristics. We incorporated modifications to traditional operant paradigms by using three video playback systems to deliver random-dot kinematogram motion stimuli coupled with salient computer-animated secondary reinforcers representative of biologically important appetitive stimuli. This procedure has the capacity to test other visual aspects in lizards as well as other non- human species using video playback and computer-animation techniques as experimental tools. Psychophysical studies that investigate the visual systems in various taxa are often challenging and require tedious trial-and-error testing before establishing a successful procedure (Ozlak & Wickens, 1999; Feldman & Balch, 2004). Some protocols have already been established for successful replication on similar taxa, or can provide a basis in which to test the sensory system in completely different species. However, not all available procedures are applicable to every species. Researchers must then develop a new method using basic conditioning paradigms. Limitations are also governed by the way in which the species is motivated to respond. These problematic features can be a function of the biological nature of the * Kevin L. Woo was supported by the Centre for the Integrative Study of Animal Behaviour postgraduate award, and the Australian Research Council.