The Role of the Glycocalyx in the Pathophysiology of Subarachnoid Hemorrhage-Induced Delayed Cerebral Ischemia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vocabulario De Morfoloxía, Anatomía E Citoloxía Veterinaria
Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) Servizo de Normalización Lingüística Universidade de Santiago de Compostela COLECCIÓN VOCABULARIOS TEMÁTICOS N.º 4 SERVIZO DE NORMALIZACIÓN LINGÜÍSTICA Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) 2008 UNIVERSIDADE DE SANTIAGO DE COMPOSTELA VOCABULARIO de morfoloxía, anatomía e citoloxía veterinaria : (galego-español- inglés) / coordinador Xusto A. Rodríguez Río, Servizo de Normalización Lingüística ; autores Matilde Lombardero Fernández ... [et al.]. – Santiago de Compostela : Universidade de Santiago de Compostela, Servizo de Publicacións e Intercambio Científico, 2008. – 369 p. ; 21 cm. – (Vocabularios temáticos ; 4). - D.L. C 2458-2008. – ISBN 978-84-9887-018-3 1.Medicina �������������������������������������������������������������������������veterinaria-Diccionarios�������������������������������������������������. 2.Galego (Lingua)-Glosarios, vocabularios, etc. políglotas. I.Lombardero Fernández, Matilde. II.Rodríguez Rio, Xusto A. coord. III. Universidade de Santiago de Compostela. Servizo de Normalización Lingüística, coord. IV.Universidade de Santiago de Compostela. Servizo de Publicacións e Intercambio Científico, ed. V.Serie. 591.4(038)=699=60=20 Coordinador Xusto A. Rodríguez Río (Área de Terminoloxía. Servizo de Normalización Lingüística. Universidade de Santiago de Compostela) Autoras/res Matilde Lombardero Fernández (doutora en Veterinaria e profesora do Departamento de Anatomía e Produción Animal. -
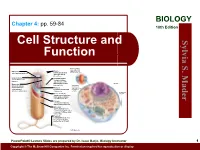
Cell Structure and Function
BIOLOGY Chapter 4: pp. 59-84 10th Edition Cell Structure and S. Sylvia Function Plasma membrane: outer surface that Ribosome: Fimbriae: regulates entrance site of protein synthesis hairlike bristles that and exit of molecules allow adhesion to Mader the surfaces Inclusion body: Conjugation pilus: stored nutrients for elongated, hollow later use appendage used for DNA transfer to other Nucleus: Mesosome: bacterial cells plasma membrane Cytoskeleton: maintains cell that folds into the Nucleoid: shape and assists cytoplasm and location of the bacterial movement of increases surface area chromosome cell parts: Endoplasmic Plasma membrane: reticulum: sheath around cytoplasm that regulates entrance and exit of molecules Cell wall: covering that supports, shapes, and protects cell Glycocalyx: gel-like coating outside cell wall; if compact, called a capsule; if diffuse, called a slime layer Flagellum: rotating filament present in some bacteria that pushes the cell forward *not in plant cells PowerPoint® Lecture Slides are prepared by Dr. Isaac Barjis, Biology Instructor 1 Copyright © The McGraw Hill Companies Inc. Permission required for reproduction or display Outline Cellular Level of Organization Cell theory Cell size Prokaryotic Cells Eukaryotic Cells Organelles Nucleus and Ribosome Endomembrane System Other Vesicles and Vacuoles Energy related organelles Cytoskeleton Centrioles, Cilia, and Flagella 2 Cell Theory Detailed study of the cell began in the 1830s A unifying concept in biology Originated from the work of biologists Schleiden and Schwann in 1838-9 States that: All organisms are composed of cells German botanist Matthais Schleiden in 1838 German zoologist Theodor Schwann in 1839 All cells come only from preexisting cells German physician Rudolph Virchow in 1850’s Cells are the smallest structural and functional unit of organisms 3 Organisms and Cells Copyright © The McGraw-Hill Companies, Inc. -

The Role of Sartans in the Treatment of Stroke and Subarachnoid Hemorrhage: a Narrative Review of Preclinical and Clinical Studies
brain sciences Review The Role of Sartans in the Treatment of Stroke and Subarachnoid Hemorrhage: A Narrative Review of Preclinical and Clinical Studies Stefan Wanderer 1,2,*, Basil E. Grüter 1,2 , Fabio Strange 1,2 , Sivani Sivanrupan 2 , Stefano Di Santo 3, Hans Rudolf Widmer 3 , Javier Fandino 1,2, Serge Marbacher 1,2 and Lukas Andereggen 1,2 1 Department of Neurosurgery, Kantonsspital Aarau, 5001 Aarau, Switzerland; [email protected] (B.E.G.); [email protected] (F.S.); [email protected] (J.F.); [email protected] (S.M.); [email protected] (L.A.) 2 Cerebrovascular Research Group, Neurosurgery, Department of BioMedical Research, University of Bern, 3008 Bern, Switzerland; [email protected] 3 Department of Neurosurgery, Neurocenter and Regenerative Neuroscience Cluster, Inselspital, Bern University Hospital, University of Bern, 3010 Bern, Switzerland; [email protected] (S.D.S.); [email protected] (H.R.W.) * Correspondence: [email protected] or [email protected]; Tel.: +41-628-384-141 Received: 17 January 2020; Accepted: 5 March 2020; Published: 7 March 2020 Abstract: Background: Delayed cerebral vasospasm (DCVS) due to aneurysmal subarachnoid hemorrhage (aSAH) and its sequela, delayed cerebral ischemia (DCI), are associated with poor functional outcome. Endothelin-1 (ET-1) is known to play a major role in mediating cerebral vasoconstriction. Angiotensin-II-type-1-receptor antagonists such as Sartans may have a beneficial effect after aSAH by reducing DCVS due to crosstalk with the endothelin system. In this review, we discuss the role of Sartans in the treatment of stroke and their potential impact in aSAH. -

Implications for Brain Injury and Protection
Antithrombotic, procoagulant, and fibrinolytic mechanisms in cerebral circulation: implications for brain injury and protection Berislav V. Zlokovic, M.D., Ph.D. Department of Neurosurgery and Division of Neurosurgery, Childrens Hospital, University of Southern California School of Medicine, Los Angeles, California Maintaining a delicate balance among anticoagulant, procoagulant, and fibrinolytic pathways in the cerebral microcirculation is of major importance for normal cerebral blood flow. Under physiological conditions and in the absence of provocative stimuli, the anticoagulant and fibrinolytic pathways prevail over procoagulant mechanisms. Blood clotting is essential to minimize bleeding and to achieve hemostasis; however, excessive clotting contributes to thrombosis and may predispose the brain to infarction and ischemic stroke. Conversely, excessive bleeding due to enhanced anticoagulatory and fibrinolytic mechanisms could predispose the brain to hemorrhagic stroke. Recent studies in the author's laboratory indicate that brain capillary endothelium in vivo produces thrombomodulin (TM), a key cofactor in the TMprotein C system that is of major biological significance to the antithrombotic properties of the blood-brain barrier (BBB). The BBB endothelium also expresses tissue plasminogen activator (tPA), a key protein in fibrinolysis, and its rapid inhibitor, plasminogen activator inhibitor (PAI-1). The procoagulant tissue factor is normally dormant at the BBB. There is a vast body of clinical evidence to document the importance of hemostasis in the pathophysiology of brain injury. In particular, functional changes caused by major stroke risk factors in the TMprotein C, tPA/PAI-1, and tissue factor systems at the BBB may result in large and debilitating infarctions following an ischemic insult. Thus, correcting this hemostatic imbalance could ameliorate drastic CBF reductions at the time of ischemic insult, ultimately resulting in brain protection. -

NIH Public Access Author Manuscript Mucosal Immunol
NIH Public Access Author Manuscript Mucosal Immunol. Author manuscript; available in PMC 2014 March 01. NIH-PA Author ManuscriptPublished NIH-PA Author Manuscript in final edited NIH-PA Author Manuscript form as: Mucosal Immunol. 2013 March ; 6(2): 379–392. doi:10.1038/mi.2012.81. Molecular Organization of the Mucins and Glycocalyx Underlying Mucus Transport Over Mucosal Surfaces of the Airways Mehmet Kesimer*,‡,▫, Camille Ehre*, Kimberly A. Burns*, C. William Davis*,†, John K. Sheehan*,‡, and Raymond J. Pickles*,§ *Cystic Fibrosis/Pulmonary Research and Treatment Center, University of North Carolina, Chapel Hill, NC 27599-7248 ‡Department of Biochemistry and Biophysics, University of North Carolina, Chapel Hill, NC 27599-7248 †Department of Cell and Molecular Physiology, University of North Carolina, Chapel Hill, NC 27599-7248 §Department of Microbiology and Immunology, University of North Carolina, Chapel Hill, NC 27599-7248 Abstract Mucus, with its burden of inspired particulates, and pathogens, is cleared from mucosal surfaces of the airways by cilia beating within the periciliary layer (PCL). The PCL is held to be ‘watery’ and free of mucus by thixotropic-like forces arising from beating cilia. With radii of gyration ~250 nm, however, polymeric mucins should reptate readily into the PCL, so we assessed the glycocalyx for barrier functions. The PCL stained negative for MUC5AC and MUC5B, but it was positive for keratan sulfate, a glycosaminoglycan commonly associated with glycoconjugates. Shotgun proteomics showed keratan sulfate-rich fractions from mucus containing abundant tethered mucins, MUC1, MUC4, and MUC16, but no proteoglycans. Immuno-histology by light and electron microscopy localized MUC1 to microvilli, MUC4 and MUC20 to cilia, and MUC16 to goblet cells. -

Unarousable Unresponsiveness in Which the Patient Lies with the Eye Closed and Has No Awareness of Self and Surroundings (2)
Neurological Disease Deborah M Stein, MD, MPH 1. Coma a. Coma is defined as “a state of extreme unresponsiveness, in which an individual exhibits no voluntary movement or behavior” (1). i. Alternatively, coma is a state of unarousable unresponsiveness in which the patient lies with the eye closed and has no awareness of self and surroundings (2). b. Coma lies on a spectrum with other alterations in consciousness – from confusion to delirium to obtundation to stupor to coma and, ultimately, brain death (2). c. To be clearly distinguished from syncope, concussion, or other states of transient unconsciousness coma must persist for at least one hour (2). d. There are 2 important characteristics of the conscious state (3) i. The level of consciousness – “arousal or wakefulness” 1. Regulated by physiological functioning and consists of more primitive responsiveness to the world such as predictable involuntary reflex responses to stimuli. 2. Arousal is maintained by the reticular activating system (RAS) - a network of structures (including the brainstem, the medulla, and the thalamus) ii. The content of consciousness – “awareness” 1. Regulated by cortical areas within the cerebral hemispheres, e. There are two main causes for coma: i. Bihemispheric diffuse cortical or white matter damage or ii. Brainstem lesions bilaterally affecting the subcortical reticular activating systems. f. A huge number of conditions can result in coma. One way to categorize these conditions is to divide them into the anatomic and the metabolic causes of coma. i. Anatomic causes of coma are those conditions that disrupt the normal physical architecture and anatomy, either at the level of the cerebral cortex or the brainstem ii. -

Nanoarchitecture and Dynamics of the Mouse Enteric Glycocalyx Examined by Freeze-Etching Electron Tomography and Intravital Microscopy
ARTICLE https://doi.org/10.1038/s42003-019-0735-5 OPEN Nanoarchitecture and dynamics of the mouse enteric glycocalyx examined by freeze-etching electron tomography and intravital microscopy Willy W. Sun1,2,5, Evan S. Krystofiak1,5, Alejandra Leo-Macias1, Runjia Cui1, Antonio Sesso3, Roberto Weigert 4, 1234567890():,; Seham Ebrahim4 & Bechara Kachar 1* The glycocalyx is a highly hydrated, glycoprotein-rich coat shrouding many eukaryotic and prokaryotic cells. The intestinal epithelial glycocalyx, comprising glycosylated transmembrane mucins, is part of the primary host-microbe interface and is essential for nutrient absorption. Its disruption has been implicated in numerous gastrointestinal diseases. Yet, due to chal- lenges in preserving and visualizing its native organization, glycocalyx structure-function relationships remain unclear. Here, we characterize the nanoarchitecture of the murine enteric glycocalyx using freeze-etching and electron tomography. Micrometer-long mucin filaments emerge from microvillar-tips and, through zigzagged lateral interactions form a three-dimensional columnar network with a 30 nm mesh. Filament-termini converge into globular structures ~30 nm apart that are liquid-crystalline packed within a single plane. Finally, we assess glycocalyx deformability and porosity using intravital microscopy. We argue that the columnar network architecture and the liquid-crystalline packing of the fila- ment termini allow the glycocalyx to function as a deformable size-exclusion filter of luminal contents. 1 Laboratory of Cell Structure and Dynamics, National Institute on Deafness and Other Communication Disorders, National Institutes of Health, Bethesda, MD 20892, USA. 2 Neuroscience and Cognitive Science Program, University of Maryland, College Park, MD 20740, USA. 3 Sector of Structural Biology, Institute of Tropical Medicine, University of São Paulo, Sao Paulo, SP 05403, Brazil. -

Predicting Recurrent Stroke After Minor Stroke and Transient Ischemic Attack
Review For reprint orders, please contact [email protected] Predicting recurrent stroke after minor stroke and transient ischemic attack Expert Rev. Cardiovasc. Ther. 7(10), 1273–1281 (2009) Philippe Couillard, The risk of a subsequent stroke following an acute transient ischemic attack or minor stroke is Alexandre Y Poppe high, with 90-day risk at approximately 10%. Identification of those patients at the highest risk and Shelagh B Coutts† for recurrent stroke following a transient ischemic attack or minor stroke may allow risk-specific †Author for correspondence management strategies to be implemented, such as hospital admission with expedited work-up Department of Clinical for those at high risk and emergency room discharge for those at low risk. Predictors of recurrent Neurosciences and Radiology, stroke, including the ABCD2 score, brain imaging and the stroke mechanism, are reviewed in University of Calgary, C1261, this article, with a focus on recent literature. An emphasis is placed on the importance of early Foothills Medical Centre, imaging of the brain parenchyma (diffusion-weighted imaging) and vascular imaging to identify 1403 29th St NW, Calgary, patients at high risk for recurrence. The need for identification of the cause of the initial event, AB, T2N 2T9, Canada allowing therapies to be tailored to the individual patient, is discussed. Tel.: 1 403 944 1594 Fax: 1 403 283 2270 KEYWORDS: imaging • prevention • prognosis • recurrence • stroke • transient ischemic attack shelagh.coutts@ albertahealthservices.ca Stroke is the second leading cause of death and is present very quickly after symptom onset. This a major cause of adult disability in the world [1,2]. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -

Mechanical Characterisation of Bone Cells and Their Glycocalyx
Mechanical characterisation of bone cells and their glycocalyx a dissertation presented by Stefania Marcotti to The Department of Mechanical Engineering University of Sheffield Sheffield, United Kingdom in partial fulfilment of the requirements for the degree of Doctor of Philosophy Supervisor: Prof. Damien Lacroix Co-Supervisor: Dr. Gwendolen C Reilly November 2017 Abstract Mechanotransductionreferstotheprocessbywhichacellisabletotranslatemech- anical stimulation into biochemical signals. In bone, mechanotransduction regu- lates how cells detect environmental stimuli and use these to direct towards bone deposition or resorption. The mechanical properties of bone cells have an impact on the way mechanical stimulation is sensed, however, little evidence is available about how these properties influence mechanotransduction. The aim of the present Thesis was to quantify the mechanical properties of bone cells with a combined experimental and computational approach. Atomic force microscopy was employed to quantify the stiffness of bone cells and their gly- cocalyx. Changes in cell stiffness during osteocytogenesis were explored. Single molecule force spectroscopy of glycocalyx components was performed to evalu- ate their anchoring to the cytoskeleton. A single cell finite element model was designed to discern the contributions of sub-cellular components in response to simulated cell nano-indentation. Wide ranges of variation were found for bone cell stiffness and a method was proposed to determine suitable sample sizes to capture population heterogeneity. Bytargetingsinglecomponentsoftheboneglycocalyx, itwaspossibletohypothes- ise different mechanotransduction mechanisms depending on the hyaluronic acid attachment to the cytoskeleton. The developed computational framework showed similar results to the nano-indentation experiments and highlighted the role of the actin cytoskeleton in withstanding compression and distributing strain within the cell. iii Publications and conference presentations Book chapters Marcotti S and Reilly GC (2017). -

Prevalence and Characteristics of Migraine in CADASIL
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Universität München: Elektronischen Publikationen Original Article Cephalalgia 2016, Vol. 36(11) 1038–1047 ! International Headache Society 2015 Prevalence and characteristics Reprints and permissions: sagepub.co.uk/journalsPermissions.nav of migraine in CADASIL DOI: 10.1177/0333102415620909 cep.sagepub.com Stephanie Guey1,2,Je´roˆme Mawet1,3, Dominique Herve´ 1,2, Marco Duering4, Ophelia Godin1, Eric Jouvent1,2, Christian Opherk5, Nassira Alili1, Martin Dichgans4,6 and Hugues Chabriat1,2 Abstract Background and objective: Migraine with aura (MA) is a major symptom of cerebral autosomal dominant arterio- pathy with subcortical infarcts and leukoencephalopathy (CADASIL). We assessed the spectrum of migraine symptoms and their potential correlates in a large prospective cohort of CADASIL individuals. Methods: A standardized questionnaire was used in 378 CADASIL patients for assessing headache symptoms, trigger factors, age at first attack, frequency of attacks and associated symptoms. MRI lesions and brain atrophy were quantified. Results: A total of 54.5% of individuals had a history of migraine, mostly MA in 84% of them; 62.4% of individuals with MA were women and age at onset of MA was lower in women than in men. Atypical aura symptoms were experienced by 59.3% of individuals with MA, and for 19.7% of patients with MA the aura was never accompanied by headache. MA was the inaugural manifestation in 41% of symptomatic patients and an isolated symptom in 12.1% of individuals. Slightly higher MMSE and MDRS scores and lower Rankin score were detected in the MA group. -

Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx
biology Article Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx Shailey Gale Twamley 1,2, Anke Stach 1, Heike Heilmann 3, Berit Söhl-Kielczynski 4, Verena Stangl 1,2, Antje Ludwig 1,2,*,† and Agnieszka Münster-Wandowski 3,*,† 1 Medizinische Klinik für Kardiologie und Angiologie, Charité—Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin, and Berlin Institute of Health, 10117 Berlin, Germany; [email protected] (S.G.T.); [email protected] (A.S.); [email protected] (V.S.) 2 DZHK (German Centre for Cardiovascular Research), Partner Site, 10117 Berlin, Germany 3 Institute of Integrative Neuroanatomy, Charité—Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin, and Berlin Institute of Health, 10117 Berlin, Germany; [email protected] 4 Institute for Integrative Neurophysiology—Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin, and Berlin Institute of Health, 10117 Berlin, Germany; [email protected] * Correspondence: [email protected] (A.L.); [email protected] (A.M.-W.) † These authors equally contributed. Simple Summary: The glycocalyx (GCX) is a hydrated, gel-like layer of biological macromolecules attached to the cell membrane. The GCX acts as a barrier and regulates the entry of external substances into the cell. The function of the GCX is highly dependent on its structure and composition. Citation: Twamley, S.G.; Stach, A.; Pathogenic factors can affect the protective structure of the GCX. We know very little about the three- Heilmann, H.; Söhl-Kielczynski, B.; dimensional organization of the GXC. The tiny and delicate structures of the GCX are difficult Stangl, V.; Ludwig, A.; to study by microscopic techniques.