Therapeutic Targeting of the General RNA Polymerase II Transcription Machinery
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

SMARCB1/INI1 Genetic Inactivation Is Responsible for Tumorigenic Properties of Epithelioid Sarcoma Cell Line VAESBJ
Published OnlineFirst April 10, 2013; DOI: 10.1158/1535-7163.MCT-13-0005 Molecular Cancer Cancer Therapeutics Insights Therapeutics SMARCB1/INI1 Genetic Inactivation Is Responsible for Tumorigenic Properties of Epithelioid Sarcoma Cell Line VAESBJ Monica Brenca1, Sabrina Rossi3, Erica Lorenzetto1, Elena Piccinin1, Sara Piccinin1, Francesca Maria Rossi2, Alberto Giuliano1, Angelo Paolo Dei Tos3, Roberta Maestro1, and Piergiorgio Modena1 Abstract Epithelioid sarcoma is a rare soft tissue neoplasm that usually arises in the distal extremities of young adults. Epithelioid sarcoma presents a high rate of recurrences and metastases and frequently poses diagnostic dilemmas. We previously reported loss of tumor suppressor SMARCB1 protein expression and SMARCB1 gene deletion in the majority of epithelioid sarcoma cases. Unfortunately, no appropriate preclinical models of such genetic alteration in epithelioid sarcoma are available. In the present report, we identified lack of SMARCB1 protein due to a homozygous deletion of exon 1 and upstream regulatory region in epithelioid sarcoma cell line VAESBJ. Restoration of SMARCB1 expression significantly affected VAESBJ cell proliferation, anchorage-independent growth, and cell migration properties, thus supporting the causative role of SMARCB1 loss in epithelioid sarcoma pathogenesis. We investigated the translational relevance of this genetic back- ground in epithelioid sarcoma and showed that SMARCB1 ectopic expression significantly augmented VAESBJ sensitivity to gamma irradiation and acted synergistically with flavopiridol treatment. In VAESBJ, both activated ERBB1/EGFR and HGFR/MET impinged on AKT and ERK phosphorylation. We showed a synergistic effect of combined inhibition of these 2 receptor tyrosine kinases using selective small-molecule inhibitors on cell proliferation. These observations provide definitive support to the role of SMARCB1 inactivation in the pathogenesis of epithelioid sarcoma and disclose novel clues to therapeutic approaches tailored to SMARCB1-negative epithelioid sarcoma. -

Human Prion-Like Proteins and Their Relevance in Disease
ADVERTIMENT. Lʼaccés als continguts dʼaquesta tesi queda condicionat a lʼacceptació de les condicions dʼús establertes per la següent llicència Creative Commons: http://cat.creativecommons.org/?page_id=184 ADVERTENCIA. El acceso a los contenidos de esta tesis queda condicionado a la aceptación de las condiciones de uso establecidas por la siguiente licencia Creative Commons: http://es.creativecommons.org/blog/licencias/ WARNING. The access to the contents of this doctoral thesis it is limited to the acceptance of the use conditions set by the following Creative Commons license: https://creativecommons.org/licenses/?lang=en Universitat Autònoma de Barcelona Departament de Bioquímica i Biologia Molecular Institut de Biotecnologia i Biomedicina HUMAN PRION-LIKE PROTEINS AND THEIR RELEVANCE IN DISEASE Doctoral thesis presented by Cristina Batlle Carreras for the degree of PhD in Biochemistry, Molecular Biology and Biomedicine from the Universitat Autònoma de Barcelona. The work described herein has been performed in the Department of Biochemistry and Molecular Biology and in the Institute of Biotechnology and Biomedicine, supervised by Prof. Salvador Ventura i Zamora. Cristina Batlle Carreras Prof. Salvador Ventura i Zamora Bellaterra, 2020 Protein Folding and Conformational Diseases Lab. This work was financed with the fellowship “Formación de Profesorado Universitario” by “Ministerio de Ciencia, Innovación y Universidades”. This work is licensed under a Creative Commons Attributions-NonCommercial-ShareAlike 4.0 (CC BY-NC- SA 4.0) International License. The extent of this license does not apply to the copyrighted publications and images reproduced with permission. (CC BY-NC-SA 4.0) Batlle, Cristina: Human prion-like proteins and their relevance in disease. Doctoral Thesis, Universitat Autònoma de Barcelona (2020) English summary ENGLISH SUMMARY Prion-like proteins have attracted significant attention in the last years. -

Integrase Interactor 1 (INI-1) Deficient Renal Cell Carcinoma
Open Access Case Report DOI: 10.7759/cureus.13082 Integrase Interactor 1 (INI-1) Deficient Renal Cell Carcinoma Manpreet Singh 1 , Harkirat Singh 1 , Benjamin Hambro 1 , Jasleen Kaur 1 , Ravi Rao 2 1. Internal Medicine, St Agnes Medical Center, Fresno, USA 2. Hematology and Oncology, St Agnes Medical Center, Fresno, USA Corresponding author: Manpreet Singh, [email protected] Abstract Members of the SWItch/sucrose nonfermentable (SWI-SNF) family, including SWI/SNF related, matrix- associated, actin-dependent regulator of chromatin, subfamily A, member 4 (SMARCA4), SWI/SNF related, matrix‐associated, actin‐dependent regulator of chromatin, subfamily B member 1 (SMARCB1)/integrase interactor 1 (INI-1) are known tumor suppressor genes. Interactions between SMARCB1/INI-1 and key protein components in various cellular pathways are related to tumor progression and proliferation. SMARCB1/INI-1 protein was undetectable in rhabdoid tumor cells, whereas non-tumorous cells express the SMARCB1/INI-1 genes. Germline and sporadic mutations of several genes encoding for proteins in this complex are known to cause a spectrum of cancers, usually with sarcomatoid features which include a very aggressive renal medullary carcinoma. We report a case of a 29-year-old male who presented with SMARCA4 deficient renal tumor with a very aggressive clinical behavior which ultimately led to his death. Categories: Nephrology, Oncology Keywords: renal medullary carcinoma, ini-1, renal cell carcinoma, swi/snf, smarcb1, integrase interactor 1 Introduction Renal medullary carcinoma is a rare and very aggressive malignancy affecting young adults with rare cases in patients with sickle cell disease or trait [1]. The tumor arises predominantly in the renal medulla and exhibits a variety of growth patterns including reticular, solid, tubular, trabecular, cribriform, sarcomatoid, and micropapillary [1]. -

Liver Med23 Ablation Improves Glucose and Lipid Metabolism Through Modulating FOXO1 Activity
Cell Research (2014) 24:1250-1265. npg © 2014 IBCB, SIBS, CAS All rights reserved 1001-0602/14 ORIGINAL ARTICLE www.nature.com/cr Liver Med23 ablation improves glucose and lipid metabolism through modulating FOXO1 activity Yajing Chu1, Leonardo Gómez Rosso1, Ping Huang2, Zhichao Wang1, Yichi Xu3, Xiao Yao1, Menghan Bao1, Jun Yan3, Haiyun Song2, Gang Wang1 1State Key Laboratory of Cell Biology, Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chi- nese Academy of Sciences, 320 Yueyang Road, Shanghai 200031, China; 2Key Laboratory of Nutrition and Metabolism, Institute for Nutritional Sciences, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, 320 Yueyang Road, Shang- hai 200031, China; 3CAS-MPG Partner Institute for Computational Biology, Shanghai Institute for Biological Sciences, Chinese Academy of Sciences, 320 Yueyang Road, Shanghai 200031, China Mediator complex is a molecular hub integrating signaling, transcription factors, and RNA polymerase II (RNAPII) machinery. Mediator MED23 is involved in adipogenesis and smooth muscle cell differentiation, suggesting its role in energy homeostasis. Here, through the generation and analysis of a liver-specific Med23-knockout mouse, we found that liver Med23 deletion improved glucose and lipid metabolism, as well as insulin responsiveness, and prevented diet-induced obesity. Remarkably, acute hepatic Med23 knockdown in db/db mice significantly improved the lipid profile and glucose tolerance. Mechanistically, MED23 participates in gluconeogenesis and cholesterol synthesis through modulating the transcriptional activity of FOXO1, a key metabolic transcription factor. Indeed, hepatic Med23 deletion impaired the Mediator and RNAPII recruitment and attenuated the expression of FOXO1 target genes. Moreover, this functional interaction between FOXO1 and MED23 is evolutionarily conserved, as the in vivo activities of dFOXO in larval fat body and in adult wing can be partially blocked by Med23 knockdown in Drosophila. -

Cyclin K Interacts with Β-Catenin to Induce Cyclin D1 Expression And
Theranostics 2020, Vol. 10, Issue 24 11144 Ivyspring International Publisher Theranostics 2020; 10(24): 11144-11158. doi: 10.7150/thno.42578 Research Paper Cyclin K interacts with β-catenin to induce Cyclin D1 expression and facilitates tumorigenesis and radioresistance in lung cancer Guojun Yao*, Jing Tang*, Xijie Yang, Ye Zhao, Rui Zhou, Rui Meng, Sheng Zhang, Xiaorong Dong, Tao Zhang, Kunyu Yang, Gang Wu and Shuangbing Xu Cancer Center, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China. *These authors contributed equally to this work. Corresponding author: Shuangbing Xu or Gang Wu, Cancer Center, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China. E-mail: [email protected] or [email protected]. © The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/). See http://ivyspring.com/terms for full terms and conditions. Received: 2019.11.29; Accepted: 2020.08.24; Published: 2020.09.11 Abstract Rationale: Radioresistance remains the major cause of local relapse and distant metastasis in lung cancer. However, the underlying molecular mechanisms remain poorly defined. This study aimed to investigate the role and regulatory mechanism of Cyclin K in lung cancer radioresistance. Methods: Expression levels of Cyclin K were measured by immunohistochemistry in human lung cancer tissues and adjacent normal lung tissues. Cell growth and proliferation, neutral comet and foci formation assays, G2/M checkpoint and a xenograft mouse model were used for functional analyses. Gene expression was examined by RNA sequencing and quantitative real-time PCR. -
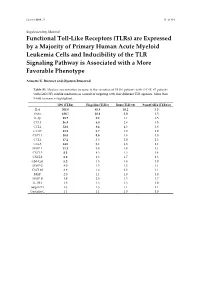
Are Expressed by a Majority of Primary Human Acute Myeloid Leukemia Cells and Inducibility of the TLR Signaling Pathway Is Associated with a More Favorable Phenotype
Cancers 2019, 11 S1 of S19 Supplementary Material Functional Toll-Like Receptors (TLRs) are Expressed by a Majority of Primary Human Acute Myeloid Leukemia Cells and Inducibility of the TLR Signaling Pathway is Associated with a More Favorable Phenotype Annette K. Brenner and Øystein Bruserud Table S1. Median concentration increase in the secretion of 19 (16 patients with G-CSF, 67 patients with GM-CSF) soluble mediators as a result of targeting with four different TLR agonists. More than 5-fold increase is highlighted. LPS (TLR4) Flagellin (TLR5) R848 (TLR7/8) Pam3CSK4 (TLR1/2) IL-6 301.0 45.9 10.2 5.3 TNFα 188.7 10.6 5.0 1.5 IL-1β 89.7 9.2 1.1 1.5 CCL3 56.4 6.0 2.4 1.5 CCL2 52.6 9.4 4.3 2.8 G-CSF 51.9 2.7 1.0 1.0 CXCL1 38.0 5.8 1.4 1.5 CCL4 17.2 3.3 2.0 1.3 CCL5 14.5 2.1 1.3 1.1 MMP-1 11.1 3.4 1.4 1.1 CXCL5 8.3 4.3 1.3 1.4 CXCL8 6.0 4.3 1.7 1.3 GM-CSF 5.2 1.5 1.4 1.0 MMP-2 4.0 1.5 1.2 1.1 CXCL10 3.9 1.6 2.2 1.1 HGF 2.0 1.1 1.0 1.0 MMP-9 1.9 2.5 1.3 1.7 IL-1RA 1.8 1.3 1.3 1.0 Serpin E1 1.3 1.3 1.1 1.1 Cystatin C 1.1 1.1 1.0 1.0 Cancers 2019, 11 S2 of S19 Table S2. -

Cytotoxic Effects and Changes in Gene Expression Profile
Toxicology in Vitro 34 (2016) 309–320 Contents lists available at ScienceDirect Toxicology in Vitro journal homepage: www.elsevier.com/locate/toxinvit Fusarium mycotoxin enniatin B: Cytotoxic effects and changes in gene expression profile Martina Jonsson a,⁎,MarikaJestoib, Minna Anthoni a, Annikki Welling a, Iida Loivamaa a, Ville Hallikainen c, Matti Kankainen d, Erik Lysøe e, Pertti Koivisto a, Kimmo Peltonen a,f a Chemistry and Toxicology Research Unit, Finnish Food Safety Authority (Evira), Mustialankatu 3, FI-00790 Helsinki, Finland b Product Safety Unit, Finnish Food Safety Authority (Evira), Mustialankatu 3, FI-00790 Helsinki, c The Finnish Forest Research Institute, Rovaniemi Unit, P.O. Box 16, FI-96301 Rovaniemi, Finland d Institute for Molecular Medicine Finland (FIMM), University of Helsinki, P.O. Box 20, FI-00014, Finland e Plant Health and Biotechnology, Norwegian Institute of Bioeconomy, Høyskoleveien 7, NO -1430 Ås, Norway f Finnish Safety and Chemicals Agency (Tukes), Opastinsilta 12 B, FI-00521 Helsinki, Finland article info abstract Article history: The mycotoxin enniatin B, a cyclic hexadepsipeptide produced by the plant pathogen Fusarium,isprevalentin Received 3 December 2015 grains and grain-based products in different geographical areas. Although enniatins have not been associated Received in revised form 5 April 2016 with toxic outbreaks, they have caused toxicity in vitro in several cell lines. In this study, the cytotoxic effects Accepted 28 April 2016 of enniatin B were assessed in relation to cellular energy metabolism, cell proliferation, and the induction of ap- Available online 6 May 2016 optosis in Balb 3T3 and HepG2 cells. The mechanism of toxicity was examined by means of whole genome ex- fi Keywords: pression pro ling of exposed rat primary hepatocytes. -

Anti-Cdk8 Antibody Produced in Rabbit (C0238)
ANTI-CDK8 Developed in Rabbit, Affinity Isolated Antibody Product Number C 0238 Product Description Reagent Anti-Cyclin-Dependent Kinase 8 (CDK8) is developed in Anti-CDK8, at approximately 1 mg/ml, is supplied as a rabbit using a synthetic peptide corresponding to the solution in phosphate buffered saline, pH 7.4 containing C-terminal region (aa 451-464) of human CDK8. The 0.2% BSA and 15 mM sodium azide. antibody is purified by protein A affinity chromatography. Anti-CDK8 specifically recognizes a 54 kDa protein Precautions and Disclaimer identified as cyclin-dependent kinase 8 (CDK8). Anti- Due to the sodium azide content, a material safety data CDK8 does not crossreact with the other members of sheet (MSDS) for this product has been sent to the CDK family. It detects human, mouse and rat CDK8. It attention of the safety officer of your institution. Consult is used in immunoblotting, immunoprecipitation and the MSDS for information regarding hazardous and safe immunofluorescence applications. handling practices. Cyclin-dependent kinase 8 (CDK8) is a 464-amino acid Storage/Stability protein containing the sequence motifs and 11 Store at –20 °C. For extended storage, upon initial subdomains characteristic of a serine/threonine-specific thawing, freeze the solution in working aliquots. kinase.1 CDKs become activated through binding to Avoid repeated freezing and thawing to prevent cyclins, formation of cyclin-CDK complexes and denaturing the antibody. Storage in “frost-free” reversible phosphorylation reactions. Cyclin-CDK freezers is also not recommended. If slight complexes directly control progression through G1, S, turbidity occurs upon prolonged storage, clarify the G2 and M phases of the cell division cycle. -

Loss of Integrase Interactor 1 (INI1) Expression in a Subset of Differentiated Thyroid Cancer
diagnostics Article Loss of Integrase Interactor 1 (INI1) Expression in a Subset of Differentiated Thyroid Cancer Kung-Chen Ho 1, Jie-Jen Lee 1, Chi-Hsin Lin 2,3, Ching-Hsiang Leung 4 and Shih-Ping Cheng 1,5,* 1 Department of Surgery, MacKay Memorial Hospital and Mackay Medical College, Taipei 104215, Taiwan; [email protected] (K.-C.H.); [email protected] (J.-J.L.) 2 Department of Medical Research, MacKay Memorial Hospital, Taipei 104215, Taiwan; [email protected] 3 Department of Bioscience Technology, Chung Yuan Christian University, Taoyuan City 320314, Taiwan 4 Division of Endocrinology and Metabolism, Department of Internal Medicine, MacKay Memorial Hospital and Mackay Medical College, Taipei 104215, Taiwan; [email protected] 5 Department of Pharmacology, School of Medicine, College of Medicine, Taipei Medical University, Taipei 110301, Taiwan * Correspondence: [email protected]; Tel.: +886-2-2543-3535 Received: 19 April 2020; Accepted: 2 May 2020; Published: 5 May 2020 Abstract: Alterations in the switching defective/sucrose non-fermenting (SWI/SNF) chromatin-remodeling complex are enriched in advanced thyroid cancer. Integrase interactor 1 (INI1), encoded by the SMARCB1 gene on the long arm of chromosome 22, is one of the core subunits of the SWI/SNF complex. INI1 immunohistochemistry is frequently used for the diagnosis of malignant rhabdoid neoplasms. In the present study, we found normal and benign thyroid tissues generally had diffusely intense nuclear immunostaining. Loss of INI1 immunohistochemical expression was observed in 8% of papillary thyroid cancer and 30% of follicular thyroid cancer. Furthermore, loss of INI1 expression was associated with extrathyroidal extension (p < 0.001) and lymph node metastasis (p = 0.038). -

Yeast Genome Gazetteer P35-65
gazetteer Metabolism 35 tRNA modification mitochondrial transport amino-acid metabolism other tRNA-transcription activities vesicular transport (Golgi network, etc.) nitrogen and sulphur metabolism mRNA synthesis peroxisomal transport nucleotide metabolism mRNA processing (splicing) vacuolar transport phosphate metabolism mRNA processing (5’-end, 3’-end processing extracellular transport carbohydrate metabolism and mRNA degradation) cellular import lipid, fatty-acid and sterol metabolism other mRNA-transcription activities other intracellular-transport activities biosynthesis of vitamins, cofactors and RNA transport prosthetic groups other transcription activities Cellular organization and biogenesis 54 ionic homeostasis organization and biogenesis of cell wall and Protein synthesis 48 plasma membrane Energy 40 ribosomal proteins organization and biogenesis of glycolysis translation (initiation,elongation and cytoskeleton gluconeogenesis termination) organization and biogenesis of endoplasmic pentose-phosphate pathway translational control reticulum and Golgi tricarboxylic-acid pathway tRNA synthetases organization and biogenesis of chromosome respiration other protein-synthesis activities structure fermentation mitochondrial organization and biogenesis metabolism of energy reserves (glycogen Protein destination 49 peroxisomal organization and biogenesis and trehalose) protein folding and stabilization endosomal organization and biogenesis other energy-generation activities protein targeting, sorting and translocation vacuolar and lysosomal -

Sudips Revised Thesis
Investigation Of The Behavior Of The Gal4 Inhibitor Gal80 Of The GAL Genetic Switch In The Yeast Saccharomyces Cerevisiae Dissertation Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Sudip Goswami, M.S. Graduate Program in Molecular Genetics The Ohio State University 2014 Dissertation Committee Dr. James Hopper, Advisor Dr. Stephen Osmani Dr. Hay-Oak Park Dr. Jian-Qiu Wu ii Copyright by Sudip Goswami 2014 iii ABSTRACT The DNA-binding transcriptional activator Gal4 and its regulators Gal80 and Gal3 constitute a galactose-responsive switch for the GAL genes of Saccharomyces cerevisiae. Gal4 binds to upstream activation sequences or UASGAL sites on GAL gene promoters as a dimer both in the absence and presence of galactose. In the absence of galactose, a Gal80 dimer binds to and masKs the Gal4 activation domain, inhibiting its activity. In the presence of galactose, Gal3 interacts with Gal80 and relieves Gal80’s inhibition of Gal4 activity allowing rapid induction of expression of GAL genes. In the first part of this work (Chapter 2) in-vitro chemical crosslinking coupled with SDS PAGE and native PAGE analysis were employed to show that the presence of Gal3 that can interact with Gal80 impairs Gal80 self association. In addition, live cell spinning disK confocal imaging showed that dissipation of newly discovered Gal80-2mYFP/2GFP clusters in galactose is dependent on Gal3’s ability to interact with Gal80. In the second part (Chapter 3), extensive analysis of Gal80 clusters was carried out which showed that these clusters associate strongly with the GAL1-10-7 locus and this association is dependent on the presence of the UASGAL sites at this locus. -

Human Cyclin-Dependent Kinase 12 (CDK12), Kinase Domain a Target Enabling Package (TEP)
Human Cyclin-Dependent Kinase 12 (CDK12), Kinase Domain A Target Enabling Package (TEP) Gene ID / UniProt ID / EC CDK12, 51755 / Q9NYV4/ 2.7.11.22, 2.7.11.23 CCNK, 8812 / O75909/ - Target Nominator Gregg Morin (UBC, Canada), Nathanael Gray (Harvard) SGC Authors Sarah E. Dixon-Clarke, Jonathan M. Elkins, Nathanael S. Gray, and Alex N. Bullock Collaborating Authors S.-W. Grace Cheng1, Tinghu Zhang2, Nicholas Kwiatkowski3, Calla M. Olson2, Brian J. Abraham3, Ann K. Greifenberg4, Scott B. Ficarro2, Yanke Liang2, Nancy M. Hannett3, Theresa Manz5, Mingfeng Hao2, Bartlomiej Bartkowiak6, Arno L. Greenleaf6, Jarrod A. Marto2, Matthias Geyer4, Richard A. Young3, and Gregg B. Morin1 Target PI Alex Bullock (SGC Oxford) Therapeutic Area(s) Oncology Disease Relevance CDK12 loss sensitises cancer cells to DNA damage Date approved by TEP 17th June 2016 Evaluation Group Document version Version 3 Document version date April 2018 DOI https://doi.org/10.5281/zenodo.1219680 Affiliations 1. Canada's Michael Smith Genome Sciences Centre, BC Cancer Agency 2. Department of Cancer Biology, Dana-Farber Cancer Institute 3. Whitehead Institute for Biomedical Research 4. Department of Structural Immunology, Institute of Innate Immunity 5. Pharmaceutical and Medicinal Chemistry, Saarland University 6. Department of Biochemistry, Duke University Medical Center USEFUL LINKS (Please note that the inclusion of links to external sites should not be taken as an endorsement of that site by the SGC in any way) SUMMARY OF PROJECT Cyclin-dependent kinase 12 (CDK12) phosphorylates RNA Pol II C-terminal domain (CTD) to promote transcriptional elongation of large DNA damage response genes. CDK12 is frequently mutated or amplified in cancer and its loss sensitises cells to DNA damage.