PDF File Created from a TIFF Image by Tiff2pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Street Tree Master Plan Report © Sunshine Coast Regional Council 2009-Current
Sunshine Coast Street Tree Master Plan 2018 Part A: Street Tree Master Plan Report © Sunshine Coast Regional Council 2009-current. Sunshine Coast Council™ is a registered trademark of Sunshine Coast Regional Council. www.sunshinecoast.qld.gov.au [email protected] T 07 5475 7272 F 07 5475 7277 Locked Bag 72 Sunshine Coast Mail Centre Qld 4560 Acknowledgements Council wishes to thank all contributors and stakeholders involved in the development of this document. Disclaimer Information contained in this document is based on available information at the time of writing. All figures and diagrams are indicative only and should be referred to as such. While the Sunshine Coast Regional Council has exercised reasonable care in preparing this document it does not warrant or represent that it is accurate or complete. Council or its officers accept no responsibility for any loss occasioned to any person acting or refraining from acting in reliance upon any material contained in this document. Foreword Here on our healthy, smart, creative Sunshine Coast we are blessed with a wonderful environment. It is central to our way of life and a major reason why our 320,000 residents choose to live here – and why we are joined by millions of visitors each year. Although our region is experiencing significant population growth, we are dedicated to not only keeping but enhancing the outstanding characteristics that make this such a special place in the world. Our trees are the lungs of the Sunshine Coast and I am delighted that council has endorsed this master plan to increase the number of street trees across our region to balance our built environment. -

Yandina Street Tree Strategy
Yandina Street tree strategy Description of area and land use Canopy cover Street tree planting strategies The local plan area of Yandina occurs in the north of the Sunshine Coast Canopy cover over all lands is below-average for the region (31%) with Street trees enhance the historical look and feel of the township and Council region and totals 396 hectares in land area. The plan area contains the Foliage and Shade Cover plan for Yandina showing that open rural reinforce existing planting themes. the Yandina township, rural residential streets, farmlands, and industrial lands account for numerous areas of low or no tree cover. Vegetation cover and commercial precincts. Originally known as 'Native Dog Flat' the oldest reported for road reserve areas is also below average (27%). Analysis of Street tree planting focuses on shading pedestrian networks, building surveyed town in the Maroochy Shire was named Yandina in 1871. street tree occupancy within the town suggests that canopy cover can be canopy and establishing feature trees in key locations; and improving the readily increased through a solid program of proactive street tree planting. Yandina's landscape character beautifully blends the cultural heritage general amenity of town approaches. values of the small country town with the natural character of the area. Major opportunities and constraints The town's strong character tree palette bleeds out into surrounding Yellow flame trees frame the distinct facade of the village shop fronts while streets and links the sports precinct and other community facilities back clumps of eucalypts grow in areas immediately surrounding the township Numerous opportunities to build on the existing street tree canopy of to the town centre with feature and shade tree plantings. -

Rdna) Organisation
OPEN Heredity (2013) 111, 23–33 & 2013 Macmillan Publishers Limited All rights reserved 0018-067X/13 www.nature.com/hdy ORIGINAL ARTICLE Dancing together and separate again: gymnosperms exhibit frequent changes of fundamental 5S and 35S rRNA gene (rDNA) organisation S Garcia1 and A Kovarˇı´k2 In higher eukaryotes, the 5S rRNA genes occur in tandem units and are arranged either separately (S-type arrangement) or linked to other repeated genes, in most cases to rDNA locus encoding 18S–5.8S–26S genes (L-type arrangement). Here we used Southern blot hybridisation, PCR and sequencing approaches to analyse genomic organisation of rRNA genes in all large gymnosperm groups, including Coniferales, Ginkgoales, Gnetales and Cycadales. The data are provided for 27 species (21 genera). The 5S units linked to the 35S rDNA units occur in some but not all Gnetales, Coniferales and in Ginkgo (B30% of the species analysed), while the remaining exhibit separate organisation. The linked 5S rRNA genes may occur as single-copy insertions or as short tandems embedded in the 26S–18S rDNA intergenic spacer (IGS). The 5S transcript may be encoded by the same (Ginkgo, Ephedra) or opposite (Podocarpus) DNA strand as the 18S–5.8S–26S genes. In addition, pseudogenised 5S copies were also found in some IGS types. Both L- and S-type units have been largely homogenised across the genomes. Phylogenetic relationships based on the comparison of 5S coding sequences suggest that the 5S genes independently inserted IGS at least three times in the course of gymnosperm evolution. Frequent transpositions and rearrangements of basic units indicate relatively relaxed selection pressures imposed on genomic organisation of 5S genes in plants. -

Native Trees and Shrubs for Seaside Areas
NATIVE TREES & SHRUBS RECOMMENDED FOR SEASIDE AREAS NOOSA & DISTRICT LANDCARE GROUP STATION STREET, POMONA PH: 5485 2468 PLANTS SUITABLE FOR SEASIDE AREAS This list has been prepared as a guide to the selection of trees and shrubs suitable for planting on the coastal areas of Queensland. Most species are available from Noosa Landcare’s nurseries. Others listed are generally available from nurseries specializing in native plants. The problems of seaside plantings are mainly concerned with salt-laden winds which burn the foliage of tender plants. For such exposed sites the following plants will provide a resistant windbreak. SPECIES COMMON NAME (SHRUBS – Up to 5 metres) Acacia podalryiifolia Queensland silver wattle Acacia spectabilis Glory or Mudgee wattle Acacia suaveolens Sweet wattle Alectryon coriaceous Beach bird’s eye Baeckea frutescens Weeping baeckea Banksia ericiifolia Heath-leaved banksia Banksia oblingifolia Dwarf banksia Banksia spinulosa var. spinulosa Spiny-leaved honeysuckle Banksia spinulosa var. collina Hair-pin banksia Callistemon citrinus Crimson bottlebrush Callistemon pachyphyllus Wallum bottlebrush Callistemon rigidus Stiff bottlebrush Grevillea banksii Bank’s grevillea Hakea sericea White hakea Leptospermum brachyandrum Weeping tea-tree Leptospermum laevigatum Coast tea-tree Leptospermum petersonii Lemon-scented tea-tree Leptospermum polygalyfolium Wild may Melaleuca armillaris Bracelet honey myrtle Melaleuca alternifolia Paperbark Melaleauca bracteata River tea-tree Melaleuca nodosa Prickly-leaved paperbark Petalostigma -

Research Article the Potential of Bunya Nut Extracts As Antibacterial Functional Food Agents
The Potential of Bunya Nut Extracts as Antibacterial Functional Food Agents Author Vesoul, J., Cock, Ian Published 2012 Journal Title Pharmacognosy Communications DOI https://doi.org/10.5530/pc.2012.1.13 Copyright Statement © 2012 Phcog.net. The attached file is reproduced here in accordance with the copyright policy of the publisher. Please refer to the journal's website for access to the definitive, published version. Downloaded from http://hdl.handle.net/10072/46944 Griffith Research Online https://research-repository.griffith.edu.au Pharmacognosy Communications www.phcogcommn.org Volume 2 | Issue 1 | Jan-Mar 2012 Research Article The Potential of Bunya Nut Extracts as Antibacterial Functional Food Agents J. Vesoulb,c I. E. Cocka,b*, aEnvironmental Futures Centre, Nathan Campus, Griffith University, 170 Kessels Rd, Nathan, Queensland 4111, Australia bBiomolecular and Physical Sciences, Nathan Campus, Griffith University, 170 Kessels Rd, Nathan, Queensland 4111, Australia cEcole Supérieure d’Ingénieurs en Développement Agroalimentaire Intégré, Université de la Réunion, Parc Technologique, 2 rue Joseph Wetzell, 27490 Sainte Clotilde, Ile de La Réunion ABSTRACT: Introduction: A.bidwillii is an endemic Australian plant traditionally used by Australian Aborigines as a food. Its nuts are known to keep well, raising the possibility that they may contain antimicrobial compounds and therefore may have value as a functional food to retard spoilage and prevent food poisoning, as well as potential medicinal antibiotic uses. Methods: A.bidwillii solvent extracts were tested for antimicrobial activity against a panel of bacteria (Aeromonas hydrophila, Alcaligenes feacalis, Bacillus cereus, Citrobacter freundii, Escherichia coli, Klebsiella pneumoniae, Proteus mirabilis, Pseudomonas fluorescens, Salmonella newport, Serratia marcescens, Shigella sonnei, Staphylococcus aureus, Staphylococcus epidermidis and Streptococcus pyogenes) and toxicity in vitro. -

Of Foresters Farmers and Fish Booklet
Of Foresters, Farmers and Fish Tales from the Wildwood of the Old and New Worlds Of Foresters, Farmers and Fish Tales from the Wildwood of the Old and New Worlds Exhibition dates Impress Printmakers Brisbane 26th November – 16th December 2019 National Botanic Garden of Wales, Aberglasney 11th April – 7th June 2020 Aberystwyth Printmakers, UK & IMPRESS Printmakers, Brisbane, Australia International Print Exchange 2019-21: An Art-Geoscience Collaborative Project To societies in the modern world, trees and woodlands have come to embody a physical, and in many cases, emotional or even spiritual link to nature. They are often used to symbolise the fragility and beauty of the natural world, and the urgency of moving to sustainable ways to live. Concepts such as human loss, growth, regeneration and renewal often use tree-like metaphors, and those responsible for the custodianship of woodlands, arboreta and botanical gardens are seen as guardians of a collective past, of the present and a hope for an increasingly uncertain future. This is further underscored by the recognition that the Earth’s forests are an important producer of oxygen and regulator of atmospheric carbon dioxide levels and that large-scale deforestation has been an important factor in the long-term warming and cooling of the planet. The perceived role that woodlands play in so-called “ecosystem services” is now seen as an integral component of the survival and quality of life on earth. Important roles include flood mitigation, soil and water conservation, carbon capture, air quality in cities, protection of biological diversity of plants and wildlife and the mental and physical wellbeing of humanity. -

CX/PR 18/50/6 February 2018 JOINT FAO/WHO FOOD STANDARDS PROGRAMME CODEX COMMITTEE on PESTICIDE RESIDUES 50Th Session Haikou, PR
E Agenda Item 7(a) CX/PR 18/50/6 February 2018 JOINT FAO/WHO FOOD STANDARDS PROGRAMME CODEX COMMITTEE ON PESTICIDE RESIDUES 50th Session Haikou, PR. China, 9 - 14 April 2018 REVISION OF THE CLASSIFICATION OF FOOD AND FEED: CLASS A: PRIMARY FOOD COMMODITIES OF PLANT ORIGIN| TYPE 04: NUTS, SEEDS AND SAPS Group 022 - Tree nuts Group 023 - Oilseeds Group 024 - Seed for beverages and sweets Group 025 - Tree saps (AT STEPS 4 AND 7) (Prepared by the Electronic Working Group chaired by the United States of America and co-chaired by the Netherlands) Codex members and Observers wishing to submit comments at Step 6 on Group 024 Seed for beverages and sweets should do so as instructed in CL 2018/12-PR. Codex members and Observers wishing to submit comments at Step 3 on Group 025 Tree saps should do so as instructed in CL 2018/13-PR. Circular letters are available on the Codex webpage/Circular Letters: http://www.fao.org/fao-who-codexalimentarius/resources/circular-letters/en BACKGROUND 1. Background on the discussion of the revision of the Classification of Food and Feed (CXM 4-1989) can be found in the reports of the 36th – 49th sessions of the Committee on Pesticide Residues (CCPR) including relevant sessions of the Codex Alimentarius (CAC) held from 2004 to 2017.1 2. The practice adopted by the Committee in the revision of the Classification is to retain the revision of the commodity groups (e.g. fruit commodity groups, vegetable commodity groups, etc.) awaiting final compilation of the related commodity groups and the corresponding tables on examples of representative commodities for inclusion in the Classification of Food and Feed and the Principles and Guidance on the Selection of Representative Commodities for the Extrapolation of Maximum Residue Limits for Pesticides to Commodity Groups (CXG 84-2012) respectively. -

Native Species Recommended for Planting As Bushtucker
NATIVE SPECIES RECOMMENDED FOR PLANTING AS BUSHTUCKER NOOSA & DISTRICT LANDCARE GROUP STATION STREET, POMONA PH: 5485 2468 ACMENA INGENS Red apple ACMENA SMITHII Creek lilly pilly ACRONYCHIA WILCOXIANA Silver aspen ALECTRYON TOMENTOSA Hairy alectryon ALPINIA CAERULEA Native ginger APHANANTHE PHILIPPINENSIS Rough-leaved elm ARAUCARIA BIDWILLII Bunya pine AUSTROMYRTUS DULCIS Midyim AUSTROMYRTUS HILLII Scaly myrtle BACKHOUSIA CITRIODORA Lemon scented myrtle BRACHYCHITON ACERIFOLIUS Flame tree CAPPARIS ARBOREA Bush caper CAPPARIS SARMENTOSA Scrambling caper CARISSA OVATA Native currant CISSUS ANTARCTICA Native grape CITRUS AUSTRALIS Round Lime DAVIDSONIA PRURIENS Davidson's plum DIANELLA CONGESTA Flax Lily DIPLOGLOTTIS AUSTRALIS Native tamarind DIPLOGLOTTIS CAMPBELLII Small leaf tamarind DRYPETES DEPLANCHEI Yellow tulip ELAEOCARPUS GRANDIS Blue quandong EUGENIA REINWARDTIANA Beach Cherry EUPOMATIA LAURINA Bolwarra EXOCARPUS CUPPRESSIFORMIS Cherry ballart EXOCARPUS LATIFOLIUS Native cherry FICUS CORONATA/FRASERI Sandpaper figs FICUS MACROPHYLLA Moreton bay fig GAHNIA ASPERA Sawsedge LOMANDRA LONGIFOLIA Matrush MACADAMIA INTEGRIFOLIA Queensland nut MACADAMIA TETRAPHYLLA Bopple nut MACLURA COCHINCHINENSIS Cockspur Thorn MELODORUM LEICHHARDTII Zig-zag, vine MISCHARYTERA LAUTERERANA Corduroy tamarind PITTOSPORUM MULTIFLORUM Orange thorn PITTOSPORUM SPINESCENS Native lime PLANCHONELLA AUSTRALIS Black apple PLEIOGYNIUM TIMORENSE Burdekin plum PODOCARPUS ELATUS Brown pine STERCULIA QUADRIFIDA Peanut tree SYZYGIUM AUSTRALE Brush cherry SYZYGIUM LUEHMANNII Riberry SYZYGIUM MOOREI Rose apple SYZYGIUM OLEOSUM Blue lilly pilly Australian native plants used as - Bush tucker food - ACACIAS — The gum of Australian species when soaked in water tends to form a jelly-like substance, which can be eaten. However, depending on the amount of tannins in the gum it may be too bitter and astringent for most palates. Pale amber gums are usually more pleasant than those that are a darker red-brown colour. -

Habitat, Population Structure and the Conservation Status of Araucaria Bidwillii Hook
ResearchOnline@JCU This file is part of the following reference: Picone, Andrew Phillip (2014) Habitat, population structure and the conservation status of Araucaria bidwillii Hook. in the Australian Wet Tropics. PhD thesis, James Cook University. Access to this file is available from: http://researchonline.jcu.edu.au/44651/ The author has certified to JCU that they have made a reasonable effort to gain permission and acknowledge the owner of any third party copyright material included in this document. If you believe that this is not the case, please contact [email protected] and quote http://researchonline.jcu.edu.au/44651/ Habitat, population structure and the conservation status of Araucaria bidwillii Hook. in the Australian Wet Tropics. Thesis submitted by Andrew Phillip Picone (18856273) October 2015 For the degree of Masters in the College of Marine and Environmental Sciences James Cook University. ACKNOWLEDGEMENTS My supervisors Professor Paul Gadek, Dr Will Edwards and Dr Charles Clarke have demonstrated great patience in my part-time progress and provide support and guidance when sought. I’m particularly grateful for Prof. Gadek’s support for the original research proposal and shared interest in the Araucariaceae. The statistical approach was greatly improved under guidance from Dr Edwards and Dr Clarke provided invaluable advice on approaching the conservation assessment. Comments on overall style and structure were gratefully received from all supervisors. I’d also like to thank the College of Marine and Environmental Sciences at James Cook University. At the Wet Tropics Management Authority, Dr Steve Goosem and Mike Stott provided advice and data on vegetation mapping for Mount Lewis. -
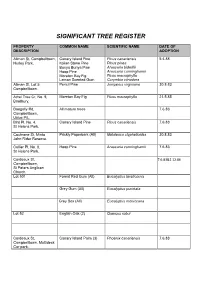
Significant Tree Register
SIGNIFICANT TREE REGISTER PROPERTY COMMON NAME SCIENTIFIC NAME DATE OF DESCRIPTION ADOPTION Allman St, Campbelltown, Canary Island Pine Pinus canariensis 5.4.88 Hurley Park. Italian Stone Pine Pinus pinea Bunya Bunya Pine Araucaria bidwillii Hoop Pine Araucaria cunninghamii Moreton Bay Fig Ficus macrophylla Lemon Scented Gum Corymbia citriodora Allman St, Lot 5, Pencil Pine Juniperus virginiana 30.8.83 Campbelltown. Athel Tree Cr, No. 9, Moreton Bay Fig Ficus macrophylla 21.5.85 Bradbury. Badgally Rd, All mature trees 7.6.83 Campbelltown, Utilux P/L. Bird Pl, No. 4, Canary Island Pine Pinus canariensis 7.6.83 St Helens Park. Cochrane St, Minto Prickly Paperbark (All) Melaleuca styphelioides 30.8.83 John Rider Reserve. Collier Pl, No. 8, Hoop Pine Araucaria cunninghamii 7.6.83 St Helens Park. Cordeaux St, 7.6.83&2.12.86 Campbelltown, St Peters Anglican Church. Lot 501 Forest Red Gum (All) Eucalyptus tereticornis Grey Gum (All) Eucalyptus punctata Grey Box (All) Eucalyptus moluccana Lot 52 English Oak (2) Quercus robur Cordeaux St, Canary Island Palm (3) Phoenix canariensis 7.6.83 Campbelltown, Multideck Car park. PROPERTY COMMON NAME SCIENTIFIC NAME DATE OF DESCRIPTION ADOPTION Cumberland Rd, Ingleburn, Cedar of Lebanon Cedrus libani 19.6.90 Hallinan Park. Cockscomb Coral Tree (2) Erythrina crista-galli Camphor Laurel Cinnamomum Prickly Paperbark camphora Melaleuca styphelioides Hansens Rd, No. 32, Camphor Laurel (2) Cinnamomum camphora 7.6.83 Minto Heights. Ingleburn Rd, Ingleburn, Bunya Bunya Pine Araucaria bidwillii 29.11.83 Ingleburn Railway Station. Kenny Cl, St Helens Park. Bunya Bunya Pine Araucaria bidwillii 7.6.83 (footpath adjacent to 24). -

Friends of the Koala Nursery
Friends of the Koala Nursery Rifle Range Road, East Lismore NSW 2480 (PO BOX 5034, East Lismore NSW 2480) * OPEN BY APPOINTMENT * Contact: Mark Wilson, Nursery Manager 0413 339 554 Email: [email protected] PLANT LIST – JUNE 2021 1. EUCALYPTS: (a) Koala food - price $1.00 (Commercial price $2.00) E. microcorys TALLOWOOD E. grandis FLOODED GUM E. robusta SWAMP MAHOGANY E. tereticornis FOREST RED GUM E. resinifera RED MAHOGANY E. siderophloia GREY IRONBARK E. saligna SYDNEY BLUE GUM E. propinqua GREY GUM E. acmenoides WHITE MAHOGANY E. dunni DUNN’S WHITE GUM E. amplifolia CABBAGE GUM E. racemosa SCRIBBLY GUM E. pilularis BLACKBUTT (b) Non-Koala food - prices as marked 2.00 Corymbia citriodora LEMON-SCENTED GUM 30m, lemon-scented foliage 2.00 Corymbia gummifera RED BLOODWOOD 30m large white flowers, good timber tree 1.50 Corymbia intermedia PINK BLOODWOOD 30m large white flowers, good timber tree 2.00 Corymbia maculata SPOTTED GUM 30m, good timber tree 2.00 Eucalyptus moluccana GREY BOX 25m mottled bark, good honey tree 2. SHRUBS: Order Price Variety Description 1.50 Acacia suaveolens SWEET-SCENTED WATTLE 1-2m, pale yellow sweetly scented flowers 3.00 Acmena ‘Allyn Magic’ DWARF LILLY-PILLY 50cm, burgundy new growth all year, 3.00 Acmena ‘Forest Flame’ 2-3m, lovely red new foliage, psyllid-free, great screen plant 3.00 Acmena smithii ‘Minipilly’ DWARF LILLY-PILLY 2m, red tips, great hedge or container plant 3.00 Astartea fascicularis ‘Pink’ 1m, pink flowers from Autumn to Summer 3.00 Austromyrtus ‘Copper Tops’ 1.2m, spreading shrub -

Two Representative Tribes of Queensland with an Inquiry Concerning the Origin of the Australian Race
f«G :REBREilSlllliE otIEKSeW'I•-:::••:•:;•••• .•• ••....:..,;.•.;.•;.;:•;.::;• %„^ ja.M:N:--M^rM:iW i9 .'^^^ THE LIBRARY , OF THE UNIVERSITY OF CALIFORNIA LOS ANGELES E hart te fau— E toro te jaaro— E nau te taata." [/'H:x-UsJ-c^ to TWO REPRESENTATIVE TRIBES OF QUEENSLAND [A// Rights Reserved^ TWO REPRESENTATIVE TRIBES OF QUEENSLAND WITH AN IN^n{r CONCE%NING THE ORIGIN OF THE AUSTRALIAN %ACE BY JOHN MATHEW, M.A., B.D. AUTHOR OF "EAGLEHAWK AND CROW," "AUSTRALIAN ECHOES," ETC. WITH AN INTRODUCTION BY Prof. A. H. KEANE, LL.D., F.R.A.I., F.R.G.S. LATE VICE-PRESIDENT R. ANTHROP. INSTITUTE AUTHOR OF "ethnology," " MAN PAST AND PRESENT," "THE WORLD'S PEOPLES," ETC. AND A MAP AND UX ILLUSTRATIONS T. FISHER UNWIN LONDON • LEIPSIC Adelphi Terrace Inselstrasse 20 1910 DEDICATED TO J. H. MACFARLAND, Esq., M.A., LL.D., MASTER OF ORMOND COLLEGE, VICE-CHANCELLOR OF MELBOURNE UNIVERSITY, AND A MEMBER OF THE BOARD FOR THE PROTECTION OF THE ABORIGINES IN THE STATE OF VICTORIA, AS A MARK OF ESTEEM AND A TOKEN OF APPRECIATION OF HIS SERVICES TO THE CAUSE OF LEARNING. CONTENTS PACE CHAP. - Introduction - - - xi - Preface . - - xxi I. Inquiry concerning the Origin of the Australian Race 25 II. The Country of the Kabi and Wakka Tribes - - - - - <i) III. Physical and Mental Characters 72 — — —Clothing - IV. Daily Life Shelter Food ^ V. Man-Making and Other Ceremonies VI. Disease and Treatment—Death—Burial and Mourning 1 10 VII. Art — Implements — Utensils —Weapons —Corroborees VIII. Social Organisation IX. The Family—Kinship and Marriage X. Religion and Magic XI.