Ir a Tabla De Contenido
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Os Nomes Galegos Dos Moluscos
A Chave Os nomes galegos dos moluscos 2017 Citación recomendada / Recommended citation: A Chave (2017): Nomes galegos dos moluscos recomendados pola Chave. http://www.achave.gal/wp-content/uploads/achave_osnomesgalegosdos_moluscos.pdf 1 Notas introdutorias O que contén este documento Neste documento fornécense denominacións para as especies de moluscos galegos (e) ou europeos, e tamén para algunhas das especies exóticas máis coñecidas (xeralmente no ámbito divulgativo, por causa do seu interese científico ou económico, ou por seren moi comúns noutras áreas xeográficas). En total, achéganse nomes galegos para 534 especies de moluscos. A estrutura En primeiro lugar preséntase unha clasificación taxonómica que considera as clases, ordes, superfamilias e familias de moluscos. Aquí apúntase, de maneira xeral, os nomes dos moluscos que hai en cada familia. A seguir vén o corpo do documento, onde se indica, especie por especie, alén do nome científico, os nomes galegos e ingleses de cada molusco (nalgún caso, tamén, o nome xenérico para un grupo deles). Ao final inclúese unha listaxe de referencias bibliográficas que foron utilizadas para a elaboración do presente documento. Nalgunhas desas referencias recolléronse ou propuxéronse nomes galegos para os moluscos, quer xenéricos quer específicos. Outras referencias achegan nomes para os moluscos noutras linguas, que tamén foron tidos en conta. Alén diso, inclúense algunhas fontes básicas a respecto da metodoloxía e dos criterios terminolóxicos empregados. 2 Tratamento terminolóxico De modo moi resumido, traballouse nas seguintes liñas e cos seguintes criterios: En primeiro lugar, aprofundouse no acervo lingüístico galego. A respecto dos nomes dos moluscos, a lingua galega é riquísima e dispomos dunha chea de nomes, tanto específicos (que designan un único animal) como xenéricos (que designan varios animais parecidos). -

A Novel Interaction: the Thin Stripe Hermit Crab, Clibanarius
A NOVEL INTERACTION: THE THIN STRIPE HERMIT CRAB, CLIBANARIUS VITTATUS, KILLS THE FLORIDA CROWN CONCH, MELONGENA CORONA, FOR ITS SHELL by Jennifer Cutter A Thesis Submitted to the Faculty of Charles E. Schmidt College of Science In Partial Fulfillment of the Requirements for the Degree of Master of Science Florida Atlantic University Boca Raton, FL August 2017 Copyright by Jennifer Cutter 2017 ii ACKNOWLEDGEMENTS I would like to thank Florida Atlantic University, Harbor Branch Oceanographic Institute, and Dr. Donna Devlin for giving me the opportunity to conduct this fascinating study. I would also like to thank the other committee members (Dr. Vincent Encomio, Dr. Edward Proffitt, and Dr. William Brooks) for their help, advice, and guidance. This work was made possible through funding from the Indian River Lagoon Research Fellowship awarded by the Harbor Branch Foundation and a scholarship awarded by The Broward Shell Club. Additionally, I would like to thank Dr. Richard Turner for being willing to meet with me on several occasions to answer questions and share his vast knowledge. iv ABSTRACT Author: Jennifer Cutter Title: A Novel Interaction: The thin stripe hermit Crab, Clibanarius vittatus, kills the Florida crown conch, Melongena corona, for its shell Institution: Florida Atlantic University Thesis Advisor: Dr. Donna Devlin Degree: Master of Science Year: 2017 The hermit crab Clibanarius vittatus kills Melongena corona solely to acquire a better fitting shell. This finding is contrary to previous studies, which found that hermit crabs of other species cannot kill gastropods or, in most instances, remove freshly dead gastropods from their shells. This interaction cannot be classified as predation because Melongena tissue was never consumed. -

Impacto De La Depredación Por Melongena Melongena (L.) Sobre Las Poblaciones Del Ostión Cmssostren Virginica (Gmelin) En La Laguna De Tampamachoco, Veracruz
http://dx.doi.org/10.7773/cm.v15i2.635 IMPACTO DE LA DEPREDACION POR Melongena melongena (L.) SOBRE LAS POBLACIONES DEL OSTION Crassostrea virginica (Gmelin) EN LA LAGUNA DE TAMPAMACHOCO, VERACRUZ IMPACT OF PREDATION BY Melongena melongena (L.) ON THE POPULATIONS OF THE OYSTER Crussosfrea virginica (Gmelin) IN LAGUNA DE TAMPAMACHOCO, VERACRUZ Guillermo Villarreal Chávez Facultad de Ciencias Marinas Universidad Autónoma de Baja California Apartado Postal 453 Ensenada, Baja California, México Villarreal Chávez, G. Impacto de la depredación por Melongena melongena (L.) sobre las poblaciones del ostión Cmssostren virginica (Gmelin) en la Laguna de Tampamachoco, Veracruz. Impact of predation by Melongena melongena (L.) on the populations of the oyster Cmssostrea virginica (Gmelin) in Laguna de Tampamachoco, Veracruz. Ciencias Marinas, 15(Z): 556.5, 1989. RESUMEN Con el objetivo de determinar el grado de impacto de la depredación del caracol Melongena melongena sobre el ostión Cmwostma vitginica se realizó una investigación en la Laguna de Tampamachoco, cerca de la ciudad de Tuxpan, Veracruz. Los resultados indican que la densidad del caracol en la zona de estudio es de 0.0007 individuos/m2, mientras que si sólo consideramos los cuadrantes en la zona de distribución preferencial es de 0.00437 individuos/m2. El caracol habita en todos los sustratos muestreados, pero se encuentra preferentementeen la zona y tipo de sustrato que prefiere el ostión, y su dieta incluye en proporción apreciable a la ostra (28%). La importancia de la depredación por caracoles, como causa de muerte no es de consideración debido a que la densidad de población de Melongena melongena es muy baja. -

Dinámica Poblacional Y Reproductiva De Turbinella Angulata Y Busycon Perversum (Mesogasteropoda: Turbinellidae Y Melongenidae) En El Banco De Campeche, México
Dinámica poblacional y reproductiva de Turbinella angulata y Busycon perversum (Mesogasteropoda: Turbinellidae y Melongenidae) en el Banco de Campeche, México Josefina Santos Valencia1,2, Martha Enríquez Díaz1 & Dalila Aldana Aranda1 1. Laboratorio de Biología y Cultivo de Moluscos. Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Mérida, Carretera Antigua a Progreso, Km. 6, A.P. 73 Cordemex, C.P. 97310, Mérida, Yucatán, México; [email protected], [email protected], [email protected] 2. Centro Regional de Investigaciones Pesqueras del Instituto Nacional de Pesca, Carretera Campeche-Lerma km. 5 S/N A.P. 140, C.P. 24500, San Francisco de Campeche, Campeche, México. Recibido 01-XII-2011. Corregido 04-VI-2012. Aceptado 05-VII-2012. Abstract: Turbinella angulata and Busycon perversum (Mesogasteropoda: Turbinellidae and Melongenidae) population and reproductive dynamics in the Campeche Bank, Mexico. Turbinella angulata and Busycon perversum are two marine gastropods that support the 90% of conch fisheries in the Gulf of Mexico, neverthe- less there is no information available to support a management strategy for both species. The aim of this work was to obtain preliminary fishery information to promote management actions for both species in the Campeche Bank. The population of conchs was sampled in 12 transects (150x1.5m) by SCUBA diving from April 2008 to March 2010. All conchs were collected and siphonal length (SL) was measured. These samples were comple- mented with data from the commercial catches, analyzing a total of 2 832 individuals of T. angulata. Siphonal length varied between 63 and 282mm. Sex ratio was of 0.98:1 female:male. -

Estado Actual De Melongena Melongena (Mollusca: Gastropoda) En La Bahía De Cispatá, Caribe Colombiano
Revista MVZ Córdoba 2020; 25(2):e1873. https://doi.org/10.21897/rmvz.1873 Original Estado actual de Melongena melongena (Mollusca: Gastropoda) en la bahía de Cispatá, Caribe Colombiano Daniela Niño-Miranda1 Biol; Alejandro Córdoba-Martínez1 Biol; Luz Arias-Reyes2 Esp; Jorge Quirós-Rodríguez3* M.Sc. 1Universidad de Córdoba, Facultad de Ciencias Básicas, Departamento de Biología, Montería, Colombia. 2Universidad de Córdoba, Facultad de Medicina Veterinaria y Zootecnia, Departamento de Ciencias Acuícolas, Montería, Colombia. 3Universidad de Córdoba, Facultad de Ciencias Básicas, Departamento de Biología, Grupo de Investigación Química de los Productos Naturales: PRONAT, Montería, Colombia. *Correspondencia: [email protected] Recibido: Noviembre 2019; Aceptado: Mayo 2020; Publicado: Julio 2020. RESUMEN Objetivo. Evaluar el estado actual de Melongena melongena en la bahía de Cispatá, Caribe Colombiano a través de los aspectos poblacionales de la especie. Materiales y métodos. Se establecieron tres sectores de muestreo, conforme con la zonificación de los manglares en la bahía. Para la recolección del material biológico se ubicó un transecto lineal de 4 x 20 m perpendicular al área del manglar con dos replicas a una distancia de 50 m. Para la determinación del sexo, se tomó una submuestra de 10 individuos con tallas superiores a la estimada para la madurez sexual de la especie. A las conchas se les midió la longitud total, con el fin de diferenciar las tallas de hembras y machos. En cada sector, la temperatura del agua y salinidad fueron registradas in situ, empleando un medidor multiparámetro Extech EC170, mientras la trasparencia del agua se calculó mediante el disco Secchi. Resultados. Se registraron un total de 1.149 individuos de M. -
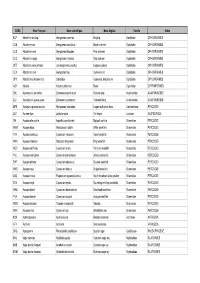
Nom Français
CODE Nom Français Nom scientifique Nom Anglais Famille Ordre KCP Abadèche du Cap Genypterus capensis Kingklip Ophidiidae OPHIDIIFORMES CUB Abadèche noir Genypterus maculatus Black cusk-eel Ophidiidae OPHIDIIFORMES CUS Abadèche rosé Genypterus blacodes Pink cusk-eel Ophidiidae OPHIDIIFORMES CUC Abadèche rouge Genypterus chilensis Red cusk-eel Ophidiidae OPHIDIIFORMES OFZ Abadèche sans jambes Lamprogrammus exutus Legless cuskeel Ophidiidae OPHIDIIFORMES CEX Abadèches nca Genypterus spp Cusk-eels nei Ophidiidae OPHIDIIFORMES OPH Abadèches, brotules nca Ophidiidae Cusk-eels, brotulas nei Ophidiidae OPHIDIIFORMES ALR Ablette Alburnus alburnus Bleak Cyprinidae CYPRINIFORMES ZML Acanthure à pierreries Zebrasoma gemmatum Spotted tang Acanthuridae ACANTHUROIDEI ZLV Acanthure à queue jaune Zebrasoma xanthurum Yellowtail tang Acanthuridae ACANTHUROIDEI MPS Achigan à grande bouche Micropterus salmoides Largemouth black bass Centrarchidae PERCOIDEI LQT Acmée râpe Lottia limatula File limpet Lottiidae GASTROPODA ISA Acoupa aile-courte Isopisthus parvipinnis Bigtooth corvina Sciaenidae PERCOIDEI WEW Acoupa blanc Atractoscion nobilis White weakfish Sciaenidae PERCOIDEI YNV Acoupa cambucu Cynoscion virescens Green weakfish Sciaenidae PERCOIDEI WKK Acoupa chasseur Macrodon ancylodon King weakfish Sciaenidae PERCOIDEI WEP Acoupa du Pérou Cynoscion analis Peruvian weakfish Sciaenidae PERCOIDEI YNJ Acoupa mongolare Cynoscion jamaicensis Jamaica weakfish Sciaenidae PERCOIDEI SWF Acoupa pintade Cynoscion nebulosus Spotted weakfish Sciaenidae PERCOIDEI WKS Acoupa -

Cercaria Criollisima Sp. N. from a Marine Snail, Melongena Melongena L., in Venezuela
OF WASHINGTON, VOLUME 39, NUMBER 2, JULY 1972 239 Khalil, L. F. 1963. On Acantlwstomum gym- Stunkard, H. W. 1931. Further observations narchi (Dollfus, 1950), with notes on the on the occurrence of anal openings in digenetic genera Acanthostomum Looss, 1899, Atropho- trematodes. Z. Parasitenk. 3: 713-725. caecum Bhalerao, 1940, Gymnatotrema Moro- . 1938. Parasitic flatworms from Yucatan. sov, 1955 and Haplocaecum Simha, 1958. J. Publ. Carneg. Inst. 491: 33-50. Helminth. 37: 207-214. Szidat, L. 1954. Trematodes nuevos de peces de Koelle, G. B., and J. S. Friedenwald. 1949. A agua dulce de la Republica Argentina. Rev. histochemical method for localizing cholin- Inst. Nac. Inves. Cienc. Nat. 3: 1-85. esterase activity. Proc. Soc. Exp. Biol. Med. Teixeira de Freitas, J. F., and H. Lent. 1938. 70: 617-622. Pesquisas helminthologicas realisadas no Mane-Garzon, F., and O. Gil. 1961. Trem- Estado do Para. II. Dois novos trematodeos atodos de las tortugas del Uruguay, II. Com. de Caiman sclerops Gray. Mem. Inst. Osw. Zool. Mus. Hist. Nat. Montevideo 5: 1-14. Cruz 33: 53-58. Morosov, H. 1955. Acanthostomatidae. In Tubangui, M. A., and V. A. Masilungan. 1936. K. I. Skrjabin (ed.), Trematodes of Animal Trematode parasites of Philippine vertebrates. and Man 10: 243-355. Moscow. VIII. Flukes from a cobra and a crocodile. Perez Vigueras, I. 1956. Contribucion al cono- Phil.J. Sci. 60: 255-266. cimiento de la fauna helmintologica cubana Yamaguti, S. 1954. Parasitic worms mainly (continuacion). Mem. Soc. Cubana Hist. Nat. from Celebes. Part 4. Trematodes of reptiles 23: 1-36. and birds. Acta Med. -

Effects of Organotins on Female Gastropods – Bibliography of Literature Read
Electronic Supplementary Material (ESI) for Journal of Environmental Monitoring This journal is © The Royal Society of Chemistry 2011 Effects of Organotins on Female Gastropods – Bibliography of Literature Read 1. Abidli, S., Lahbib, Y., and El Menif, N. T. 2009a. Imposex and genital tract malformations in Hexaplex trunculus and Bolinus brandaris collected in the Gulf of Tunis. B Mar Sci. 85: 11 - 25. 2. Abidli, S., Lahbib, Y., and El Menif, N. T. 2009b. Effects of TBT on the imposex development, reproduction and mortality in Hexaplex trunculus (Gastropoda: Muricidae). J Mar Biol Assoc UK. 89: 139 - 146. 3. Alvarez, M. M. S., and Ellis, D. V. 1990. Widespread neogastropod imposex in the northeast Pacific - Implications for TBT contamination surveys. Mar Pollut Bull. 21: 244 - 247. 4. Alzieu, C. 2000. Impact of tributyltin on marine invertebrates. Ecotoxicol. 9: 71 - 76. 5. An, W., and Hu, J. Y. 2006. Effects of endocrine disrupting chemicals on China's rivers and coastal waters. Front Ecol Environ. 4: 378 - 386. 6. Andersen, L. E. 2004a. Imposex: A biological effect of TBT contamination in Port Curtis, Queensland. Aust J Ecotoxicol. 10: 105 - 113. 7. Andersen, L. 2004b. Imposex in the City - A survey to monitor the effects of TBT contamination in Port Curtis, Queensland. Cooperative Research Centre for Coastal Zone Estuary and Waterway Management. pp. 25 pp. 8. Arconada, B., and Ramos, M. A. 2002. Spathogyna, a new genus for Valvata (? Tropidina) fezi Altimira, 1960 from eastern Spain: Another case of pseudohermaphroditism in the Hydrobiidae (Gastropoda). J Mollus Stud. 68: 319 - 327. 9. Axiak, V., Vella, A. -

East Coast Marine Shells; Descriptions of Shore Mollusks Together With
fi*": \ EAST COAST MARINE SHELLS / A • •:? e p "I have seen A curious child, who dwelt upon a tract Of Inland ground, applying to his ear The .convolutions of a smooth-lipp'd shell; To yi'hJ|3h in silence hush'd, his very soul ListehM' .Intensely and his countenance soon Brightened' with joy: for murmerings from within Were heai>^, — sonorous cadences, whereby. To his b^ief, the monitor express 'd Myster.4?>us union with its native sea." Wordsworth 11 S 6^^ r EAST COAST MARINE SHELLS Descriptions of shore mollusks together with many living below tide mark, from Maine to Texas inclusive, especially Florida With more than one thousand drawings and photographs By MAXWELL SMITH EDWARDS BROTHERS, INC. ANN ARBOR, MICHIGAN J 1937 Copyright 1937 MAXWELL SMITH PUNTZO IN D,S.A. LUhoprinted by Edwards B'olheri. Inc.. LUhtiprinters and Publishert Ann Arbor, Michigan. iQfj INTRODUCTION lilTno has not felt the urge to explore the quiet lagoon, the sandy beach, the coral reef, the Isolated sandbar, the wide muddy tidal flat, or the rock-bound coast? How many rich harvests of specimens do these yield the collector from time to time? This volume is intended to answer at least some of these questions. From the viewpoint of the biologist, artist, engineer, or craftsman, shellfish present lessons in development, construction, symme- try, harmony and color which are almost unique. To the novice an acquaint- ance with these creatures will reveal an entirely new world which, in addi- tion to affording real pleasure, will supply much of practical value. Life is indeed limitless and among the lesser animals this is particularly true. -

Catalogue of Type Specimens 4. Linnaean Specimens
Uppsala University Museum of Evolution Zoology section Catalogue of type specimens. 4. Linnaean specimens 1 UPPSALA UNIVERSITY, MUSEUM OF EVOLUTION, ZOOLOGY SECTION (UUZM) Catalogue of type specimens. 4. Linnaean specimens The UUZM catalogue of type specimens is issued in four parts: 1. C.P.Thunberg (1743-1828), Insecta 2. General zoology 3. Entomology 4. Linnaean specimens (this part) Unlike the other parts of the type catalogue this list of the Linnaean specimens is heterogenous in not being confined to a physical unit of material and in not displaying altogether specimens qualifying as types. Two kinds of links connect the specimens in the list: one is a documented curatorial tradition referring listed material to collections handled and described by Carl von Linné, the other is associated with the published references by Linné to literary or material sources for which specimens are available in the Uppsala University Zoological Museum. The establishment of material being 'Linnaean' or not (for the ultimate purpose of a typification) involves a study of the history of the collections and a scrutiny of individual specimens. An important obstacle to an unequivocal interpretation is, in many cases, the fact that Linné did not label any of the specimens included in the present 'Linnaean collection' in Uppsala (at least there are no surviving labels or inscriptions with his handwriting or referable to his own marking of specimens; a single exception will be pointed out below in the historical survey). A critical examination must thus be based on the writings of Linné, a consideration of the relation between between these writings and the material at hand, and finally a technical and archival scrutiny of the curatorial arrangements that have been made since Linné's time. -

Zoogeography of Marine Gastropod in the Southern Caribbean. a New Look
Caribbean Journal of Science, Vol. 31, No. 1-2, 104-121, 1995 Copyright 1995 College of Arts and Sciences University of Puerto Rico, Mayaguez Zoogeography of Marine Gastropod in the Southern Caribbean: A New Look at Provinciality JUAN M. DIAZ Instituto de Investigaciones Marinas de Punta de Betin -lNVEMAR-, Apartado 1016, Santa Martar Colombia ABSTRACT . – Recorded occurrences of 266 species of higher Gastropoda were arranged in nine subareas along the coastal areas of southern Central America and northern South America. The value of different gastropod families as zoogeographic indicators and the degree of faunal “singularity” of subareas were inferred respectively from the mean value of the index of “Breadth of Geographic Range” (BGR) of species involved. BGR was lower among families exhibiting predominantly direct development. About 43% of species are widely distributed in the tropical Western Atlantic, whereas approximately 40% are endemic to the southern Caribbean – enough to consider this area as a separate zoogeographic province. Northern Venezuela, Santa Marta, and the Leeward Islands are the subareas richest in species, whereas the subarea between the Orinoco delta and Surinam is the most depauperate. The number of species occurring in a given subarea is more closely related to environmental heterogeneity than to the shelf extent of the subarea. A similarity level of 50% distinguishes five zoogeographic areas within the lower Caribbean, two of them as transitional to other tropical Western Atlantic Provinces and the other three are proposed as subprov- inces. Trade wind-induced upwelling along the coasts of northern Colombia and Venezuela on the one hand, and zoogeographic links of the present molluscan fauna to the Eastern Pacific on the other, are the main factors explaining the present distribution patterns of marine gastropod in the southern Caribbean. -

Were Directly Com- Pared with Three Specimens (Cl. 13 to 15 Mm) from Tortugas, Florida (July 1925, H
CRUSTACEA DECAPODA OF SURINAME 121 between 4 and 8 mm (6 mm in the ovigerous female), were directly com- pared with three specimens (cl. 13 to 15 mm) from Tortugas, Florida (July 1925, H. Boschma; Leiden Mus. Reg. No. Crust. D 12587). The differences between the Suriname and Florida specimens have already been mentioned in the above description. They are so slight that, for the time being at least, the specimens are considered conspecific. In Schmitt's (1935) drawing of this species the spinules on the antennae and the walking legs are not shown, while also the characteristic squamiform sculpturation of these legs is omitted. Type locality. Off Fort Jefferson dock, Garden Key, Dry Tortugas, Florida, U.S.A.; interstices of large Porites clumps. Distribution. The species was only known from Florida (U.S.A.) and Puerto Rico; it is now reported for the first time from Suriname. Paguristes oxyophthalmus new species (textfigs. 22b, 23) Coquette Investigations About 20 miles N.N.W. of the mouth of the Coppename River; depth 31 m; 1-5 April 1957; first voyage. — 1 ovigerous female. (L) N.N.W. of the mouth of the Marowijne River, about 20 miles offshore; depth 27 m; 29 April-3 May 1957; fifth voyage. — 1 male. (L) Station 20, N.E. of the mouth of the Suriname River, 6° 28' N 54° 57.5' W; bottom shells; depth 31 m; 11 May 1957. — 1 male. (L) Station 28, N.E. of the mouth of the Suriname River, 6° 48'N 54°54'W; bottom shells; depth 46 m; 12 May 1957.