Archivio Istituzionale Open Access Dell'università Di Torino Occurrence and Impact of Agrilus Spp. And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

International Poplar Commission Poplars, Willows and People's Wellbeing
INTERNATIONAL POPLAR COMMISSION 23rd Session Beijing, China, 27 – 30 October 2008 POPLARS, WILLOWS AND PEOPLE’S WELLBEING Synthesis of Country Progress Reports Activities Related to Poplar and Willow Cultivation and Utilization, 2004 through 2007 October 2008 Forest Resources Development Service Working Paper IPC/6E Forest Management Division FAO, Rome, Italy Forestry Department Disclaimer Nineteen member countries of the IPC have provided national progress reports to the 23rd Session of the International Poplar Commission. A Synthesis has been made by the Food and Agriculture Organization of the United Nations (FAO) and summarizes issues, highlights status and identifies trends affecting cultivation, management and utilization of Poplars and Willows in temperate and boreal regions of the world. Comments and feedback are welcome. For further information, please contact: Mr. Jim Carle Secretary International Poplar Commission Forestry Department Food and Agriculture Organization of the United Nations (FAO) Viale delle Terme di Caracalla I-00153 Rome ITALY E-mail: [email protected] For quotation: FAO, October 2008. Synthesis of Country Progress Reports received, prepared for the 23rd Session of the International Poplar Commission, jointly hosted by FAO and by the Beijing Forestry University, the State Forest Administration of China and the Chinese Academy of Forestry; Beijing, China, 27-30 October 2008. International Poplar Commission, Working, Paper IPC/6. Forest Management Division, FAO, Rome (unpublished). Web references: For details relating to the International Poplar Commission as a Technical Statutory Body of FAO, including National Poplar Commissions, working parties and initiatives, can be viewed on www.fao.org/forestry/ipc, and highlights of the 23rd Session of the International Poplar Commission 2008 can be viewed on www.fao.org/forestry/ipc2008. -

DGC 39 Del 13/05/2020
COPIA ALBO COMUNE DI DIANO D’ALBA PROVINCIA DI CUNEO VERBALE DI DELIBERAZIONE DELLA GIUNTA COMUNALE N.39 del 13/05/2020 OGGETTO: CUP: D27E20000010007. BANDO "INTERVENTI FARO" : LANGA DEL SOLE. RESTAURO CONSERVATIVO DI SPIANAMENTO - SAN SEBASTIANO DI DIANO D'ALBA E SUA VALORIZZAZIONE NEL SISTEMA TURISTICO E CICLO-TURISTICO DI LANGA E DELL'ALTA LANGA - FASE IV. SOTTOPROGETTO 2 : I. A. N.U.A. ALLESTIMENTO DEL SISTEMA INFORMATICO E DELLE FINESTRE PANORAMA - 2° LOTTO FUNZIONALE (ALBARETTO DELLA TORRE - CERRETTO LANGHE - SERRAVALLE LANGHE). APPROVAZIONE PROGETTO DEFINITIVO L’anno duemilaventi, addì tredici, del mese di maggio alle ore 19:00 nella solita sala delle riunioni previo esaurimento delle formalità prescritte dall’art. 27 dello Statuto Comunale vennero per oggi convocati i componenti di questa Giunta Comunale. All’appello risultano: Cognome e Nome Carica Presente Assente CARDINALE Ezio Sindaco X RINALDI Sergio Vice Sindaco X DESTEFANIS Fabrizio Assessore X TARICCO Cristina Assessore X GHISOLFI Sara Assessore X Totale Presenti: 4 Totale Assenti: 1 Assiste alla seduta il Segretario Comunale Dott. Maurizio Rita SCUNCIO, il quale provvede alla redazione del presente verbale. Documento prodotto con sistema automatizzato del Comune di Diano d'Alba. (D.Lgs. n. 39/93 art.3). La presente copia è destinata unicamente alla pubblicazione sull'albo pretorio on-line Essendo legale il numero degli intervenuti, CARDINALE Ezio nella sua qualità di Sindaco, riconosciuta legale l’adunanza, assume la presidenza e dichiara aperta la seduta per la trattazione dell’oggetto sopra indicato. Documento prodotto con sistema automatizzato del Comune di Diano d'Alba. (D.Lgs. n. 39/93 art.3). -

International Poplar Commission
INTERNATIONAL POPLAR COMMISSION 25th Session Berlin, Germany, 13- 16 September 2016 Poplars and Other Fast-Growing Trees - Renewable Resources for Future Green Economies Synthesis of Country Progress Reports - Activities Related to Poplar and Willow Cultivation and Utilization- 2012 through 2016 September 2016 Forestry Policy and Resources Division Working Paper IPC/15 Forestry Department FAO, Rome, Italy Disclaimer Twenty-one member countries of the IPC, and Moldova, the Russian Federation and Serbia, three non-member countries, have provided national progress reports to the 25th Session of the International Poplar Commission. A synthesis has been made by the Food and Agriculture Organization of the United Nations that summarizes issues, highlights status and identifies trends affecting the cultivation, management and utilization of poplars and willows in temperate and boreal regions of the world. Comments and feedback are welcome. For further information, please contact: Mr. Walter Kollert Secretary International Poplar Commission Forestry Department Food and Agriculture Organization of the United Nations (FAO) Viale delle Terme di Caracalla 1 I-00153 Rome Italy E-mail: [email protected] For quotation: FAO, 2016. Poplars and Other Fast-Growing Trees - Renewable Resources for Future Green Economies. Synthesis of Country Progress Reports. 25th Session of the International Poplar Commission, Berlin, Federal Republic of Germany, 13-16 September 2016. Working Paper IPC/15. Forestry Policy and Resources Division, FAO, Rome. http://www.fao.org/forestry/ipc2016/en/. -

Case Study Italy
TOWN Small and medium sized towns in their functional territorial context Applied Research 2013/1/23 Case Study Report | Italy Version 15/February/2014 ESPON 2013 1 This report presents the interim results of an Applied Research Project conducted within the framework of the ESPON 2013 Programme, partly financed by the European Regional Development Fund. The partnership behind the ESPON Programme consists of the EU Commission and the Member States of the EU27, plus Iceland, Liechtenstein, Norway and Switzerland. Each partner is represented in the ESPON Monitoring Committee. This report does not necessarily reflect the opinion of the members of the Monitoring Committee. Information on the ESPON Programme and projects can be found on www.espon.eu The web site provides the possibility to download and examine the most recent documents produced by finalised and ongoing ESPON projects. This basic report exists only in an electronic version. © ESPON & University of Leuven, 2013. Printing, reproduction or quotation is authorised provided the source is acknowledged and a copy is forwarded to the ESPON Coordination Unit in Luxembourg. ESPON 2013 2 List of authors Cristiana Cabodi (Officina Territorio) Alberta de Luca (Officina Territorio) Alberto Di Gioia (Politecnico di Torino) Alessia Toldo (Officina Territorio) ESPON 2013 3 Table of contents 1. National context (p. 9) 1.1 Semantic point of view (p. 9) 1.2 Institutional and administrative point of view (p. 9) 1.3 statistical point of view (p. 9) 1.4 Small and Medium Sized Towns (SMSTs) in National/Regional settlement system: literary overview (p 12.) 1.4.1 The Italian urban system from the 1960s to the 1970s: the polarized structure (p. -

STATUS PROGETTO FWA Aggiornato Il 4 Novembre 2019 Da Infratel Italia
STATUS PROGETTO FWA aggiornato il 4 novembre 2019 da Infratel Italia Regione Provincia Comune STATUS PROGETTO FWA al 04 Novembre 2019 Piemonte Cuneo Acceglio Approvato progetto definitivo Piemonte Cuneo Aisone Approvato progetto definitivo Piemonte Cuneo Alba Approvato progetto definitivo Piemonte Cuneo Albaretto della Torre Approvato progetto definitivo Piemonte Cuneo Alto Approvato progetto definitivo Piemonte Cuneo Argentera In progettazione definitiva Piemonte Cuneo Arguello In approvazione progetto definitivo Piemonte Cuneo Bagnasco Approvato progetto definitivo Piemonte Cuneo Bagnolo Piemonte Approvato progetto definitivo Piemonte Cuneo Baldissero d'Alba Approvato progetto definitivo Piemonte Cuneo Barbaresco Approvato progetto definitivo Piemonte Cuneo Barge In esecuzione Piemonte Cuneo Barolo Approvato progetto definitivo Piemonte Cuneo Bastia Mondovì Approvato progetto definitivo Piemonte Cuneo Battifollo Approvato progetto definitivo Piemonte Cuneo Beinette Approvato progetto definitivo Piemonte Cuneo Bellino In esecuzione Piemonte Cuneo Belvedere Langhe Approvato progetto definitivo Piemonte Cuneo Bene Vagienna Approvato progetto definitivo Piemonte Cuneo Benevello Approvato progetto definitivo Piemonte Cuneo Bergolo In progettazione definitiva Piemonte Cuneo Bernezzo Approvato progetto definitivo Piemonte Cuneo Bonvicino In progettazione definitiva Piemonte Cuneo Borgo San Dalmazzo Lavori Completati Piemonte Cuneo Borgomale In approvazione progetto definitivo Piemonte Cuneo Bosia In progettazione definitiva Piemonte Cuneo -

Localita' Cap Provincia Abetone Cutigliano 51024 Pt
Elenco CAP/Località Periferiche aggiornato al 14 gennaio 2019 L'elenco che segue può subire variazioni. Si consiglia di scaricarlo una volta al mese. LOCALITA' CAP PROVINCIA ABETONE CUTIGLIANO 51024 PT ABRIOLA 85010 PZ ACCEGLIO 12021 CN ACCUMOLI 02011 RI ACERNO 84042 SA ACQUARIA 41025 MO AGAGGIO INFERIORE 18010 IM AGRA 21010 VA AIRETTA 28827 VB AIROLE 18030 IM AISONE 12010 CN ALA DI STURA 10070 TO ALAGNA VALSESIA 13021 VC ALBANETO 02016 RI ALBARETO 43051 PR ALBARETTO DELLA TORRE 12050 CN ALBERA LIGURE 15060 AL ALBERONA 71031 FG ALBOGASIO 22010 CO ALBUGNANO 14022 AT ALFERO 47028 FC ALGUA 24010 BG ALICE SUPERIORE 10010 TO ALLEGREZZE 16049 GE ALPEPIANA 16048 GE ALPICELLA 17019 SV ALTAGNANA 54100 MS ALTARE 17041 SV ALTIPIANI DI ARCINAZZO 00020 RM ALTO 12070 CN ALTO RENO TERME 40046 BO ALTO SERMENZA 13029 VC AMBORZASCO 16049 GE ANDONNO 12010 CN ANDRATE 10010 TO ANGROGNA 10060 TO APRICALE 18035 IM AQUILA DI ARROSCIA 18020 IM ARAMENGO 14020 AT ARCINAZZO ROMANO 00020 RM AREMOGNA 67037 AQ ARGENTERA 12010 CN ARGUELLO 12050 CN ARINA 32033 BL ARMO 18026 IM ARNASCO 17032 SV ARSIERO 36011 VI ARZENO D'ONEGLIA 18022 IM ASIAGO 36012 VI ASTA NELL'EMILIA 42030 RE ATELETA 67030 AQ AURIGO 18020 IM AVAGLIO 51010 PT AVENALE 62011 MC AVERARA 24010 BG AZZANO 55047 LU BACENO 28861 VB BADALUCCO 18010 IM BADIA PRATAGLIA 52014 AR BADIA TEDALDA 52032 AR BAGNARIA 27050 PV BAGNASCO 12071 CN BAGNI DI LUCCA 55022 LU BAGNI DI LUCCA VILLA 55022 LU BAGNI DI VINADIO 12010 CN BAGNO DI ROMAGNA 47021 FC BAISO 42031 RE BAJARDO 18031 IM BALESTRINO 17020 SV BALME 10070 TO BALMUCCIA -
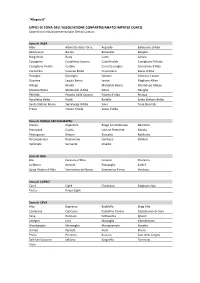
Zone Del Sistema Confartigianato Cuneo -> Comuni
“Allegato B” UFFICI DI ZONA DELL’ASSOCIAZIONE CONFARTIGIANATO IMPRESE CUNEO Zone e loro limitazione territoriale. Elenco Comuni. Zona di ALBA Alba Albaretto della Torre Arguello Baldissero d’Alba Barbaresco Barolo Benevello Bergolo Borgomale Bosia Camo Canale Castagnito Castelletto Uzzone Castellinaldo Castiglione Falletto Castiglione Tinella Castino Cerretto Langhe Corneliano d’Alba Cortemilia Cossano Belbo Cravanzana Diano d’Alba Feisoglio Gorzegno Govone Grinzane Cavour Guarene Lequio Berria Levice Magliano Alfieri Mango Montà Montaldo Roero Montelupo Albese Monteu Roero Monticello d’Alba Neive Neviglie Perletto Pezzolo Valle Uzzone Piobesi d’Alba Priocca Rocchetta Belbo Roddi Rodello Santo Stefano Belbo Santo Stefano Roero Serralunga d’Alba Sinio Tone Bormida Treiso Trezzo Tinella Vezza d’Alba Zona di BORGO SAN DALMAZZO Aisone Argentera Borgo San Dalmazzo Demonte Entracque Gaiola Limone Piemonte Moiola Pietraporzio Rittana Roaschia Robilante Roccasparvera Roccavione Sambuco Valdieri Valloriate Vernante Vinadio Zona di BRA Bra Ceresole d’Alba Cervere Cherasco La Morra Narzole Pocapaglia Sanfrè Santa Vittoria d’Alba Sommariva del Bosco Sommariva Perno Verduno Zona di CARRÙ Carrù Cigliè Clavesana Magliano Alpi Piozzo Rocca Cigliè Zona di CEVA Alto Bagnasco Battifollo Briga Alta Camerana Caprauna Castellino Tanaro Castelnuovo di Ceva Ceva Garessio Gottasecca Igliano Lesegno Lisio Marsaglia Mombarcaro Mombasiglio Monesiglio Montezemolo Nucetto Ormea Paroldo Perlo Priero Priola Prunetto Roascio Sale delle Langhe Sale San Giovanni Saliceto -

2011 Grapevine Newsletter 1.Pdf
T O O RDER M ORE F EA T URED W INES C ALL 1-800-823-5527 T ODAY ! Volume 19 Number 1 ©Vinesse Wine Club 2011 The SKU 17124 GrThe Officiala NewslepTTer fOrevine ViiNesse wiiNe club MeMbers Exploring Italy’s Langhe Valley How to Organize Your Home Wine Collection The Grades of Beef and Their Wine Equivalents Style: Why Real Maple Syrup Is So Expensive G E T Y OUR D AILY D OSE O F W INE N E W S A T www .V INESSE T ODAY . C O M 2 • The Grapevine • Visit vinesse.com OUR MISSION: To uncover and bring you wine gems diTOr s OurNal from around the world, which you’re e ’ J not likely to discover on your own, and which enhance your wine enjoyment. Steaking Claim to the YOUR GRAPEVINE TEAM: Intrepid Wine Enthusiast, Chief Best Grades of Beef Taster and Winehound: Martin Stewart By Robert Johnson Chief Operating Officer (aka “The Buck Stops Here”): he type of beef Lawrence D. Dutra T that a restaurant Editor: serves can make all the Robert Johnson difference between a Wine Steward: memorable meal and a Katie Montgomery dining disaster. The Grapevine is published by Vinesse, 822 Hampshire Road, Suite E, Westlake Beef is graded by the United States Village, CA 91361; 1-800-823-5527. Department of Agriculture, and as Copyright by Vinesse, LLC. Printed in USA. No responsibility is assumed for loss or is the case with school work, grades damage to unsolicited materials. matter. The higher the grade, the better the beef. -

Legenda T Braglia Lesegno T .! Torelli O Casc
1:70.000 Ü Presidio del Territorio PIANO FAUNISTICO VENATORIO PROVINCIALE 2003-2008 Legge 11 febbraio 1992, n. 157 articolo 10 Legge regionale 4 settembre 1996, n. 70 articolo 6 Deliberazione del Consiglio Provinciale n. 10-32 del 30 giugno 2003 Deliberazione della Giunta Regionale n. 102-10160 del 28 luglio 2003 Deliberazione della Giunta Provinciale n. 105 del 24.03.2009 e n. 47 del 30.04.2012 Deliberazione della Giunta Provinciale n. 20 del 04/05/2018 Starderi Base cartografica: CTR numerica 1/10.000 - Regione Piemonte - Settore Cartografico (autorizzazione n. 6/2002). Manzotti Cartografia ed elaborazioni GIS:Provincia di Cuneo - Settore Presidio del Territorio Pelizzeri ([email protected]) Balluri ZRC - Castiglione - Ettari 163,582011 Corso Nizza 21 – 12100 CUNEO http://www.provincia.cuneo.it/tutela_fauna/index.jsp Serra Grilli Coazzolo Neive .! Castiglione Tinella Rivetti .! Borgonuovo Serra Boella San Carlo Stazione Bric San Cristoforo Cotta Moniprandi ZRC - Valdivilla - Ettari 140,719273 Moretta Casasse Valdivilla ATC CN3 Bricco di Neive Santo Stefano Belbo ROERO .! Robini Ferrere San Maurizio ZRC - Santo Stefano - Ettari 157,953696 a CA CN1 l ATC CN2 Macarini l VALLE PO SALUZZO - SAVIGLIANO e n i T Riforno Domere ATC CN4 Giacosa S. Libera e CA CN2 ALBA - DOGLIANI t n Camo VALLE VARAITA ! e . r r Neviglie Macchia ATC CN5 o .! San Adriano Casc. Monsignore Treiso T ATC CN1 CORTEMILIA .! CA CN3 CUNEO - FOSSANO Mango VALLI MAIRA E GRANA .! Mad. della Rovere CA CN4 Trezzo Tinella .! Pianella VALLE STURA CA CN6 Leomonte VALLI MONREGALESI Meruzzano CA CN5 Naranzana VALLI GESSO, VERMENAGNA E PESIO ZRC - Cossano - Ettari 217,055045 CA CN7 .! ALTA VALLE TANARO La Cappelleta Casc. -

Codice A16000 D.D. 26 Aprile 2018, N. 154 Elenco Dei Comuni Che Hanno
REGIONE PIEMONTE BU22 31/05/2018 Codice A16000 D.D. 26 aprile 2018, n. 154 Elenco dei Comuni che hanno istituito la Commissione locale per il paesaggio ai sensi dell'articolo 4, comma 6, della legge regionale 1 dicembre 2008, n. 32 e s.m.i., per l'esercizio della funzione autorizzatoria in materia di paesaggio. (omissis) IL DIRETTORE (omissis) determina • di dare atto che il Comune sotto elencato ha assolto alle condizioni richieste dal Codice e dalla Legge regionale 1° dicembre 2008, n. 32 e s.m.i., per esercitare la funzione autorizzatoria in materia di paesaggio: Comune Prov. Provvedimento di nomina della Commissione Caramagna Piemonte CN D.G.C. di Sommariva del Bosco (Comune capofila) n. 25 del 10.02.2015* * Istituzione in forma associata della Commissione Locale per il Paesaggio • di dare atto che i Comuni sotto elencati, a seguito del rinnovo o della nuova nomina dei componenti della Commissione locale per il paesaggio, o dell’approvazione di una nuova convenzione per la gestione in forma associata della stessa Commissione, rispettano le condizioni richieste dal Codice e dalla Legge regionale 1° dicembre 2008, n. 32 e s.m.i., per esercitare la funzione autorizzatoria in materia di paesaggio, e che pertanto, il Provvedimento di nomina della Commissione deve intendersi così sostituito: Sommariva del Bosco CN D.G.C. di Sommariva del Bosco (Comune capofila) n. 25 del 10.02.2015* San Sebastiano Curone AL D.C.C. di Tortona (Comune capofila) n. 47 del 08.09.2014* * Istituzione in forma associata della Commissione Locale per il Paesaggio • di dare atto che i Comuni sotto elencati, a seguito della sostituzione di uno o più membri della Commissione locale per il paesaggio con nomina di nuovi componenti, rispettano le condizioni richieste dal Codice e dalla Legge regionale 1° dicembre 2008, n. -

All a Plessi Sottodimensionati
Allegato A - Plessi in deroga A.S. 2020 - 2021 Piano dimensionamento rete scolastica - Plessi siti in Comuni marginali e montani da mantenere in deroga ENTE PROVINCIA DI CUNEO Allegato a Deliberazione Consiglio n. del DATI A.S. 2019/20 PUNTI DI EROGAZIONE DEL SERVIZIO DA MANTENERE IN DEROGA motivazione cod MIUR Comune sede cod MIUR Grado Comune sede (comune Istituto Tipo e denominazone Indirizzo istituto istituto plesso plesso Denominazione Indirizzo plesso plesso n. marginale/monta autonomo istituto autonomo autonomo autonomo scolastico scolastico plesso scolastico scolastico scolastico alunni no) montano/alternanza CNIC81000D I.C. di Robilante Piazza della Pace, 8 Robilante CNEE81005Q Primaria di Entracque Via A. Barale 2 Entracque 31 con Valdieri Piazza della Resistenza montano/alternanza CNIC81000D I.C. di Robilante Piazza della Pace, 8 Robilante CNEE81006R Primaria di Valdieri Valdieri 24 1 con Entracque CNIC80300A I.C. L. Romano Via Peano, 6 Demonte CNAA803039 Infanzia di Vinadio Via Trocello 10 Vinadio 15 montano CNIC80300A I.C. L. Romano Via Peano, 7 Demonte CNEE80305L Primaria di Vinadio/Aisone Via trocello, 12 Vinadio 31 montano Via Rocca de'Baldi, Secondaria n allievi in aumento dopo CNIC847004 I.C. Borgo San Giuseppe Cuneo CNMM847026 di Castelletto Stura Via Cuneo, 2 Castelletto Stura 24 15 di I grado accorp a Cuneo P.zza Battaglione montano CNIC82800P I.C. G. Giolitti Dronero CNEE828051 Primaria di Prazzo Via Nazionale 22 Prazzo 12 Alpini alta marginalità P.zza Battaglione S.Damiano CNIC82800P I.C. G. Giolitti Dronero CNEE828062 Primaria di S.Damiano Macra Via A. Beltricco 5 22 montano Alpini Macra P.zza Battaglione Secondaria CNIC82800P Giovanni Giolitti Dronero CNMM82802R di Stroppo Fraz. -

Oup Foresj Cpz024 1..10 ++
Forestry An International Journal of Forest Research Forestry 2019; 00,1–10, doi:10.1093/forestry/cpz024 An illustrated guide to distinguish emerald ash borer (Agrilus planipennis) from its congeners in Europe Downloaded from https://academic.oup.com/forestry/advance-article-abstract/doi/10.1093/forestry/cpz024/5486413 by guest on 08 May 2019 Mark G. Volkovitsh1, Marina J. Orlova-Bienkowskaja2*, Alexey V. Kovalev3 and Andrzej O. Bieńkowski2 1Zoological Institute, Russian Academy of Sciences, St. Petersburg 199034, Russia 2A.N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Moscow 119071, Russia 3All-Russian Institute of Plant Protection of the Russian Academy of Agricultural Sciences, 3 Podbelsky Shosse, St. Petersburg–Pushkin 196608, Russia *Corresponding author E-mail: [email protected] Received 23 September 2018 Emerald ash borer (EAB) Agrilus planipennis is native to East Asia and has recently become a devastating alien pest of ash trees in North America and European Russia. There is no doubt that the pest will spread to other European countries. Early detection and identification of this damaging pest is crucial for minimizing its poten- tial negative economic and ecological impacts. There are about 87 species of the genus Agrilus in Europe, but there are no guides for identification, that include Agrilus planipennis and native species. We present here the first guide to distinguish emerald ash borer from similar native European Agrilus species based on easily detec- tible external morphological characters suitable for use by non-taxonomists. The main diagnostic characters of EAB adults are: large size (12–15 mm), head and pronotal disc deeply impressed, pronotal sides converging toward anterior margin, elytra mostly brightly emerald, without tomentose spots, pygidium bearing apical process.