Morphological Characterization of Wild Populations of Solanum Lycopersicum Var
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mapa De Veracruz De Ignacio De La Llave. División Municipal
Veracruz de Ignacio de la Llave N O E 1 S 203 161 150 102 114 183 150154 060 155 150 109 129 063035 151 042 T1amaulipas50 013 192 056 153 023 078 154 055 034 197 150 056 055 167 010 163 095 060 096 057 187106 50 107 156 002 133 058 177 036 166 013 1213 51 160 009 189 086 194 132 112 152 136 093 53 027 026 076 001182 087 004 034 128 205 121 202 083 157 033 025 092 065 7 175 038 San Luis Potosí 170 161 150 180 025164 154 079 088 134 150 131 046 017 060 024 155 063 150 198 129 035 013 071521 069 056 153 162 078 040 1 055 034 056 051 124 188 160 055 167 158 179 189 066 146 058 037 160 103 064 211 071 043 Hidalgo 189 067 050 008 14 027 203 029 076 047 165 200 102 114 180680 007 202 083 157 183 062 175 033 127 170 180 109 101 022 044 131 042 192 068 021 125 072 198 023 081 157 00340 3 069 124 Golfo de México 197 138118 085 196 051 158 010 163 099 101595135 014 175 066 074 113 053 031 037 030 187001986 057 185 041 052 067103 064 211 106 140098 050 107 156 002 203 102 114 177 036 166 104709 168 086 194 112 117 183 132 006 137 131 136020693 020 201 109 042 192 023 001182 087 195 001471 197 128 184 010 163 095 110 173 096 057 092 065 191 061987106 025 019 156 107 002166 038 159 040 177 036 009 164 086 194 112 025 016 132 093 079 088 134 136026 046 024 017 051 124 128 001182 087 004 150658 191 162 025 092 126 038 188 193 079025164 088 134 016 146 179 193 066 046 024 017 028 037 162 126 071 043 188 193 100 146 179 193 008 148 064 028 029 103 071 024311 047 080 165 200 067 008 148 100 186 007 090 050 029 047 080 165 200 127 062 186 007 090 Escala -

Maltrata 2240 Atanacio Lopez Moreno 21 Camerino Z
COMITÉ DIRECTIVO ESTATAL DE VERACRUZ SECRETARÍA DE ORGANIZACIÓN PRESIDENTES DE COMITÉS SECCIONALES SECCION DISTRITO MUNICIPIO NOMBRE DEL PRESIDENTE ELECTORAL 21 CAMERINO Z. MENDOZA 100 MALTRATA 2240 ATANACIO LOPEZ MORENO 21 CAMERINO Z. MENDOZA 100 MALTRATA 2241 JAIME NICOLAS PONCIANO SOLIS 21 CAMERINO Z. MENDOZA 100 MALTRATA 2242 REY DAVID HUERTA CARRERA 21 CAMERINO Z. MENDOZA 100 MALTRATA 2243 CARLOS EUFEMIO ZAMORA GONZALEZ 21 CAMERINO Z. MENDOZA 100 MALTRATA 2244 MARIA DE LA CRUZ MENDEZ VICENTE 21 CAMERINO Z. MENDOZA 100 MALTRATA 2245 ENRIQUE GUZMAN REYES 21 CAMERINO Z. MENDOZA 100 MALTRATA 2246 GERARDO JUSTINO MORALES ROMERO 21 CAMERINO Z. MENDOZA 100 MALTRATA 2247 ENRIQUE SANCHEZ MARTINEZ 21 CAMERINO Z. MENDOZA 100 MALTRATA 2248 JUAN AVELINO ROMERO COMITÉ DIRECTIVO ESTATAL DE VERACRUZ SECRETARÍA DE ORGANIZACIÓN INTEGRANTES DE LOS CONSEJOS POLÍTICOS MUNICIPALES DISTRITO MUNICIPIO NOMBRE DEL CONSEJERO 21 CAMERINO Z. MENDOZA 100 MALTRATA ALVAREZ OREA DELFINA COMITÉ DIRECTIVO ESTATAL DE VERACRUZ 21 CAMERINO Z. MENDOZA 100 MALTRATA AMADOR ONTIVEROS LOREYLA COMISIÓN ESTATAL DE PROCESOS INTERNOS 21 CAMERINO Z. MENDOZA 100 MALTRATA ANTONIO BORJA MARIBEL 21 CAMERINO Z. MENDOZA 100 MALTRATA ANTONIO HUERTA YURIANA PRESIDENTES DE COMITÉS SECCIONALES 21 CAMERINO Z. MENDOZA 100 MALTRATA ARENAS LOPEZ CRISTIAN 21 CAMERINO Z. MENDOZA 100 MALTRATA ARGUELLES GONZALEZ CIRILO FELIMON 21 CAMERINO Z. MENDOZA 100 MALTRATA AVELINO AGUILAR TERESA 21 CAMERINO Z. MENDOZA 100 MALTRATA BAROJAS CAMPOS ANTONIA GUILLERMINA 21 CAMERINO Z. MENDOZA 100 MALTRATA BELTRAN PEREZ MARINA 21 CAMERINO Z. MENDOZA 100 MALTRATA BERMUDEZ ROMERO FELIPE MIGUEL 21 CAMERINO Z. MENDOZA 100 MALTRATA BERMUDEZ VELASCO TANIA CECILIA 21 CAMERINO Z. MENDOZA 100 MALTRATA CALVARIO BERMUDEZ NARCISO ARMANDO 21 CAMERINO Z. -

Pobreza Y Rezago Social En El Estado De Veracruz
Informe Especial POBREZA Y REZAGO SOCIAL EN EL ESTADO DE VERACRUZ 1 INFORME ESPECIAL: POBREZA Y REZAGO SOCIAL EN EL ESTADO DE VERACRUZ CUENTA PÚBLICA 2018 ÍNDICE Hidalgotitlán ....................................................................................................................................................... 7 Huatusco ......................................................................................................................................................... 13 Huayacocotla ................................................................................................................................................... 19 Hueyapan de Ocampo .................................................................................................................................... 25 Huiloapan de Cuauhtémoc .............................................................................................................................. 31 Ignacio de la Llave .......................................................................................................................................... 37 Ilamatlán .......................................................................................................................................................... 43 Isla ................................................................................................................................................................. 49 Ixcatepec ........................................................................................................................................................ -
Veracruz.Pdf
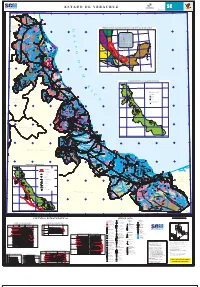
94 34’ 22 30’ 94 00’ 30’ 95 00’ 30’ 96 00’ 30’ 97 00’ 30’ 98 00’ 98 42’ 30’ 22 30’ ESTADO DE Qhoal TAMAULIPAS L. LAS PINTAS L. QUINTERO Qhoal L. JOPOY R. Tamesi Qhoal TpaeLu- Ar- Mg . L. LA TORTUGA Qhoal TpaeLu- Ar- Mg L. MAYORAZGO Puente Chairel C. LA TORTUGA A. CORCOVADO - CACALILAO Qhoal TeMg- Lu KcmLu- Mg TomAr- Lu- Lm LAGUNA TpaeLu- Ar- Mg TeMg- Lu EL CHAREL ToAr- Lu- Cgp PLANTA MET` LICA Qholi COMPA ˝A M INERA AUTL` N S.A. DE C.V. UNIDAD TAMOS EL MORALILLO PERSEVERANCIA Qhoal AutlÆn PRIMERO DE MAYO CONGLOMERADO Tamos CHAPOPOTERA ARENISCA Puente Tampico L. CHILA (GRAVA) Qhoal (ARENA, GRAVA) TomAr- Lu- Lm AN` HUAC Qhoal TeMg- Lu TpaeLu- Ar- Mg TomAr- Lu- Lm . CIUDAD CUAHUT MOC Puente PÆnuco TpaeLu- Ar- Mg ARENISCA 87 Qhoal (ARENA, GRAVA) L. TACAT` 90 22 00’ ALUVI N LAGUNA 93 (GRAVA) Qhoeo CONGLOMERADO TomAr- Lu- Lm PUEBLO VIEJO 96 (GRAVA) CONGLOMERADO Mata 99 (GRAVA) de ChÆvez ESTADO DE L. TANCHICU˝N L. LAS OLAS TeMg- Lu CARGADURA DEL GOLFO S.A. DE C.V. TeMg- Lu TAMPICO ALTO G SELECCIONADORA TpaeLu- Ar- Mg CONGLOMERADO DE MATERIAL (GRAVA) CONGLOMERADO (GRAVA) CONGLOMERADO ToAr- Lu- Cgp SAN LUIS Qhoal (GRAVA) TomAr- Lu- Lm 24 R . Qhoeo P P` NUCO Æn uco Qhoal Qhoal a p il To R. R. P TpaeLu- Ar- Mg L E Y E N D A a COA itØ POTOS˝ Qhola L. LAGARTERO Qhoeo 22 00’ TomAr- Lu- Lm Qhoal ` REA MINERALIZADA LLANO DE BUSTOS C. TRES MORILLOS L. -

Saberes Territoriales Y Manejo Campesino Del Monte
UNIVERSIDAD AUTÓNOMA METROPOLITANA UNIDAD – XOCHIMILCO DIVISIÓN DE CIENCIAS SOCIALES Y HUMANIDADES POSGRADO EN DESARROLLO RURAL NIVEL MAESTRIA EL CAMPESINADO ZONGOLIQUEÑO EN LA DISPUTA TERRITORIAL Saberes territoriales y manejo campesino del monte PRESENTA ROSALINDA HIDALGO LEDESMA DIRECTORA: DRA. ALEJANDRA TOSCANA APARICIO MÉXICO, D.F. ABRIL DE 2016 Índice Resumen ........................................................................................................................ 4 Agradecimientos ........................................................................................................... 7 Siglas.............................................................................................................................. 8 Introducción................................................................................................................. 11 Antecedentes y justificación ...................................................................................... 11 Objetivos ...................................................................................................................... 14 Apuestas epistemológicas y herramental teórico .................................................... 16 Estrategia metodológica ............................................................................................. 27 1. Los bosques, sus pobladores y los contextos de la política forestal en México................................................................................................................ 29 1.1. Los -

Stratigraphy of the 8.5 – 9.0 Ka BP Citlaltépetl Pumice Fallout Sequence
Revista Mexicana de Ciencias StratigraphyGeológicas, v.of 21, the núm. 8.5 – 3, 9.0 2004, ka B.P. p. 353-370 Citlaltépetl pumice fallout sequence 353 Stratigraphy of the 8.5 – 9.0 ka B.P. Citlaltépetl pumice fallout sequence Andrea Rossotti1,*, Gerardo Carrasco-Núñez2 1 Posgrado en Ciencias de la Tierra 2 Centro de Geociencias Universidad Nacional Autónoma de México, Campus Juriquilla, A.P. 1–742, 76001 Querétaro, Qro., Mexico. * [email protected] ABSTRACT Citlaltépetl is an active volcano presently in a dormant state, belonging to the easternmost part of the Mexican Volcanic Belt. Although the volcano is characterized by long periods of volcanic repose, some explosive plinian eruptions occurred in Holocene time. In particular, an eruptive period between 8.5–9.0 ka B.P. originated an alternated sequence of pumice and scoria flows called Citlaltépetl Ignimbrite (0.26 km3 DRE) and a few ash and lapilli fallout deposits (Citlaltépetl Pumice) with a wide dispersion range around the cone. In this work we present a detailed reconstruction of the fallout Citlaltépetl Pumice stratigraphy based on the combined study of 107 vertical sections, grain-size and component analysis of each layer. The eruptive sequence comprises a succession of pyroclastic deposits, including four major eruptive episodes. New radiocarbon dating on charcoal fragments interbedded in the flow deposits beneath the fallout and in the fallout itself show ages younger than 9,475±160 yr. B.P. and older than 8,505±50 yr. B.P., respectively, which is in accordance with previous dates. Stratigraphy and dating allowed a precise stratigraphic correlation of the most representative fallout deposits of the Holocene history of Citlaltépetl volcano and their relationship with associated pyroclastic flows. -

Descriptivo De La Distritacion Federal Veracruz Marzo 2017
DESCRIPTIVO DE LA DISTRITACION FEDERAL VERACRUZ MARZO 2017 DESCRIPTIVO DE LA DISTRITACION FEDERAL VERACRUZ REGISTRO FEDERAL DE ELECTORES Página 1 de 21 La entidad federativa de Veracruz se integra por 20 demarcaciones territoriales distritales electorales federales, conforme a la siguiente descripción: Distrito 01 Esta demarcación territorial distrital federal tiene su cabecera ubicada en la localidad PANUCO perteneciente al municipio PANUCO, asimismo, se integra por un total de 13 municipios, que son los siguientes: NARANJOS AMATLAN, integrado por 19 secciones: de la 0320 a la 0338. CITLALTEPETL, integrado por 9 secciones: de la 0695 a la 0703. CHINAMPA DE GOROSTIZA, integrado por 10 secciones: de la 1403 a la 1412. OZULUAMA, integrado por 29 secciones: de la 2776 a la 2804. PANUCO, integrado por 83 secciones: de la 2814 a la 2889 y de la 2891 a la 2897. PUEBLO VIEJO, integrado por 40 secciones: de la 3219 a la 3258. TAMALIN, integrado por 8 secciones: de la 3543 a la 3550. TAMIAHUA, integrado por 21 secciones: de la 3551 a la 3571. TAMPICO ALTO, integrado por 19 secciones: de la 3572 a la 3590. TANCOCO, integrado por 7 secciones: de la 3591 a la 3597. TANTIMA, integrado por 11 secciones: de la 3598 a la 3608. TEMPOAL, integrado por 30 secciones: de la 3706 a la 3735. REGISTRO FEDERAL DE ELECTORES Página 2 de 21 EL HIGO, integrado por 16 secciones: de la 4696 a la 4711. El distrito electoral 01 se conforma por un total de 302 secciones electorales. Distrito 02 Esta demarcación territorial distrital federal tiene su cabecera ubicada en la localidad TANTOYUCA perteneciente al municipio TANTOYUCA, asimismo, se integra por un total de 15 municipios, que son los siguientes: BENITO JUAREZ, integrado por 10 secciones: de la 0482 a la 0491. -

Ley Orgánica Del Municipio Libre, Promulgada El Nueve De Febrero De Mil Novecientos Ochenta Y Cuatro
LEY ORGANICA DEL MUNICIPIO LIBRE ÚLTIMA REFORMA PUBLICADA EN LA GACETA OFICIAL: 10 DE AGOSTO DE 2015 Ley publicada en la Gaceta Oficial. Órgano del Gobierno del Estado de Veracruz-Llave, el viernes 5 de enero de 2001. Al margen un sello que dice: Estados Unidos Mexicanos.-Gobernador del Estado de Veracruz- Llave. Miguel Alemán Velazco, Gobernador del Estado Libre y Soberano de Veracruz-Llave, a sus habitantes sabed: Que la Honorable Quincuagésima Novena Legislatura del Congreso del Estado se ha servido dirigirme la siguiente Ley para su promulgación y publicación: Al margen un sello con el Escudo Nacional que dice: Estado Unidos Mexicanos.-Poder Legislativo.-Estado Libre y Soberano de Veracruz-Llave. La Honorable Quincuagésima Novena Legislatura del Congreso del Estado Libre y Soberano de Veracruz-Llave, en uso de la facultad que le confieren los artículos 33 fracciones I y VIII de la Constitución Política local; 47 de la Ley Orgánica del Poder Legislativo; 103 del Reglamento para el Gobierno Interior del Poder Legislativo; y en nombre del pueblo, expide la siguiente: L E Y Número 9 ORGÁNICA DEL MUNICIPIO LIBRE TÍTULO PRIMERO DISPOSICIONES GENERALES CAPÍTULO I DE LOS MUNICIPIOS DEL ESTADO Artículo 1. La presente ley tiene por objeto desarrollar las disposiciones constitucionales relativas a la organización y funcionamiento del Municipio Libre. Artículo 2. El Municipio Libre es la base de la división territorial y de la organización política y administrativa del Estado. El Municipio Libre contará con personalidad jurídica y patrimonio propios, será gobernado por un Ayuntamiento y no existirá autoridad intermedia entre éste y el Gobierno del Estado. -

Descripción De La Problemática Parque Nacional Pico De Orizaba I
Descripción de la Problemática Parque Nacional Pico de Orizaba I. Antecedentes El 4 de enero de 1937 se publicó en el Diario Oficial de la Federación el Decreto del Ejecutivo Federal mediante el cual se declara Parque Nacional El Pico de Orizaba, área destinada a la conservación perpetua de la flora y fauna comarcanas dentro de los límites siguientes: “Del Norte, partiendo de las cumbres del cerro del Río Valiente, el lindero sigue hacia el Sureste en línea recta hasta llegar al punto conocido con el nombre de Potrero Nuevo; de este lugar, el lindero sigue al Suroeste pasando por las cumbres del cerro de Palo Gacho y de Tepala o Piedras Blancas, hasta llegar al pueblo de Texmalaquilla; de este punto, el lindero voltea hacia el Noroeste hasta llegar a las inmediaciones del pueblo de Xepestepec, de donde, con dirección hacia el Noreste, el lindero termina en las cumbres del cerro del Río Valiente que se tomó como punto de partida”1 Se localiza en los municipios de Chalchicomula de Sesma, Atzizintla, Tlachichuca en Puebla y, Calcahualco y La Perla en el estado de Veracruz; entre las cotas altitudinales 3,038 a 5,636 msnm. Tienen una superficie de 19,750.005000 hectáreas, integrada por un polígono general.2 Figura 1. Volcán Pico de Orizaba o Citlaltépetl dentro del PN Pico de Orizaba. Fuente: http://portal.veracruz.gob.mx/portal/page?_pageid=313,4544264&_dad=portal&_schema=PORTAL 1 Artículo Primero del Decreto que declara Parque Nacional El Pico de Orizaba. Diario Oficial de la Federación, 4 de enero de 1937. -

Download Brochure
Structural Features Eagle Ford Oil & Gas Windows F F GeoEdges Inc. F F F F F F F F F F Texas MEXICO GEOLOGICAL EDGE SET ( ( F Sabinas Basin for all digital mapping applications F ( ( ( ( ( Mexico ( ( ( ( Parras( Basin ( ( Burgos Basin ( ( ( ( ( ( ( ( ( ( ( ( F ( Magiscatzin Basin ( ( ( ( Tampico Basin Mexico ( ( ( ( ( ( F ( Tuxpan Platform Pimienta Oil & Gas Windows( El Abra / Tamabra Facies Burgos Basin ( F ( ( F F ( F ( ( ( ( Texas( ( ( F ( Sabinas Basin ( ( ( ( Tuxpan Platform Slope ( GeoEdges Inc. provides detailed and accurate ( ( ( ( subsurface geology at your fingertips in ArcGIS, ( ( GeoGraphix, Petra and all other ESRI Shapefile ( Slope ( compatible mapping applications ( ( ( ( ( ( Parras Basin ( ( Burgos Basin ( ( ( ( ( ( ( ( Includes geological edges for Burgos, Sabinas, Tampico, Veracruz, Macuspana, and Jurassic ( ( Salt Basins, Tuxpan Platform, and Villahermosa ( ( Uplift F ( ( Includes field outlines, structural features, Slope Magiscatzin Basin outcrops, facies and reservoir trends, structure ( Platform ( and isopach maps, shale gas trends, and other ( ( resource plays ( ( Tampico Basin ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( ANNUAL LICENSE SUBSCRIPTION - 2021 Pricing Oil and Gas Fields Basins & Uplifts The Mexico Geological Edge Set is available to Mississippi subscribing companies on an annual license basis Florida Louisiana Alabama (per company, based on daily worldwide production): Texas El Burro Uplift <2,000 BOE/d USD $5,600 per year <10,000 BOE/d USD $8,000 per year <20,000 BOE/d USD $10,400 per year Sabinas -

Tlacotalpan Veracruz
TLACOTALPAN VERACRUZ AGENDA DE COMPETITIVIDAD DEL DESTINO TURÍSTICO TLACOTALPAN PATRIMONIO CULTURAL DE LA HUMANIDAD El Colegio de Veracruz Institución Local de Educación Superior (ILES) Coordinadora General Coordinadora de Destino Dra. María de los Ángeles Piñar Álvarez Lic. Esveyde del Castillo Jiménez 1 DIRECTORIO SECRETARÍA DE TURISMO FEDERAL MTRA. CLAUDIA RUIZ MASSIEU Secretaria de Turismo C. P. Carlos Manuel Joaquín González Subsecretario de Innovación y Desarrollo Turístico Lic. José Salvador Sánchez Estrada Subsecretario de Planeación Lic. Francisco Maass Peña Subsecretario de Calidad y Regulación Mtro. Octavio Mena Alarcón Oficial Mayor FONDO NACIONAL DE FOMENTO AL TURISMO LIC. HÉCTOR MARTÍN GÓMEZ BARRAZA Director General CONSEJO DE PROMOCIÓN TURÍSTICA DE MÉXICO LIC. RODOLFO LÓPEZ NEGRETE COPPEL Director General DIRECTORIO GOBIERNO DEL ESTADO DE VERACRUZ DR. JAVIER Duarte DE Ochoa Gobernador Constitucional del Estado de Veracruz Lic. Harry Grappa Guzmán Secretario de Turismo del Estado de Veracruz EL COLEGIO DE VERACRUZ Mtro. EUGENIO A. VÁSQuez MUÑoz Director General de El Colegio de Veracruz Dra. Ángeles Piñar Álvarez Directora General del Proyecto PRESENTACIÓN La prioridad del Presidente Enrique Peña Nieto ha sido emprender reformas transformadoras en los diferentes ámbitos de la vida nacional para que México sea un país en paz, incluyente, con educación de calidad, próspero y con responsabilidad global. La Política Nacional Turística tiene como objeto convertir al turismo en motor de desarrollo. Por ello trabajamos en torno a cuatro grandes directrices: ordenamiento y transformación sectorial; innovación y competitividad; fomento y promoción; y sustentabilidad y beneficio social para promover un mayor flujo de turistas y fomentar la atracción de inversiones que generen empleos y procuren el desarrollo regional y comunitario. -

Proceso Electoral Interno 2020 - 2021
PROCESO ELECTORAL INTERNO 2020 - 2021 FECHA I DE FEBRERO DE 2021 ACUERDO COEEVERJAYTOS/PRCDNC/055 ru r I A r'rftiuI*uAP.I I f'TftAI TATAI r IVPIUII1 &%I I '('%I J I i I JI_ VERACRUZ, MEDIANTE EL CUAL SE DECLARA LA PROCEDENCIA DE REGISTRO DE PRECANDIDATURA A LA PLANILLA ENCABEZADA POR EL C.MARIA DOLORES GAMBOA AGUIRRE, PARA EL AYUNTAMIENTO DE TLACOTALPAN, VERACRUZ, CON MOTIVO DEL PROCESO INTERNO DE SELECCION DE CANDIDATURAS A AYUNTAMIENTOS DEL ESTADO DE VERACRUZ, QUE REGISTRARA EL PARTIDO ACCION NACIONAL, DENTRO DEL PROCESO ELECTORAL LOCAL 2020-2021. EL PLENO DE LA COMISION ELECTORAL ESTATAL DEL PARTIDO ACCION NACIONAL EN VERACRUZ, EN RELACION CON EL PROCESO INTERNO DE SELECCION DE CANDIDATO A LAS ALCALDIAS ACAJETE, ACAYUCAN, ACTOPAN, AGUA DULCE, ALAMO TEMAPACHE, ALPATLAHUA, ALTO LUCERO, ALTOTONGA, ALVARADO, AMATLAN, ANGEL R. CABADA, ANTIGUA LA, ATOYAC, ATZACAN, ATZALAN, BANDERILLA, BENITO JUAREZ, BOCA DEL RIO, CAMERINO Z. MENDOZA, CAMARON DE TEJEDA, CARLOS A. CARR!LLO, CARRILLO PUERTO, CASTILLO DE TEAYO, CATEMACO, CAZONES DE HERRERA, CERRO AZUL, CHICONQUIACO, CHINAMPA DE GOROSTIZA, CHOAPAS LAS, CHOCAMAN, CHONTLA, CITLALTEPEC, COATEPEC, COATZACOALCOS, COMAPA, CORDOBA, COSAMALOAPAN, COSCOMATEPEC, COSOLEACAQUE, COTAXTLA, COXQUIHUI, CUICHAPA, CUITLAHUAC, EMILIANO ZAPATA, FILOMENO MATA, FORTIN, HIDALGOTITLAN, HUATUSCO, IXHUACAN DE LOS REYES, IXHUATLANCILLO, IXTACZOQUITLAN, JUAN RODRIGUEZ CLARA, JALTIPAN, JILOTEPEC, JOSE AZUETA, LANDERO Y COSS, LERDO DE TEJADA, MALTRATA, MANLIO FABIO ALTAMIRANO, MARTINEZ DE LA TORRE, MEDELLIN DE BRAVO, MINATITLAN,