Rapid Onsets of Warming Events Trigger Mass Mortality of Coral Reef Fish
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Emerging Risks from Marine Heat Waves
COMMENT DOI: 10.1038/s41467-018-03163-6 OPEN Emerging risks from marine heat waves Thomas L. Frölicher1,2 & Charlotte Laufkötter 1,2 Recent marine heat waves have caused devastating impacts on marine ecosystems. Sub- stantial progress in understanding past and future changes in marine heat waves and their risks for marine ecosystems is needed to predict how marine systems, and the goods and 1234567890():,; services they provide, will evolve in the future. Extreme climate and weather events shape the structure of terrestrial biological systems and affect the biogeochemical functions and services they provide for society in a fundamental manner1. There is overwhelming evidence that atmospheric heat waves over land are changing under global warming, increasing the risk of severe, pervasive and in some cases irreversible impacts on natural and socio-economic systems2. In contrast, we know little how extreme events in the ocean, especially those associated with warming will change under global warming, and how they will impact marine organisms. This knowledge gap is of particular concern as some of the recent observed marine heat waves (MHWs) demonstrated the high vulnerability of marine organisms and ecosystems services to such extreme climate events. Definition, observations, and key processes A marine heat wave is usually defined as a coherent area of extreme warm sea surface tem- perature (SST) that persists for days to months3. MHWs have been observed in all major ocean basins over the recent decade, but only a few MHWs have been documented and analyzed extensively (Fig. 1). One of the first MHW that has been characterized in the literature occurred in 2003 in the northwestern Mediterranean Sea with SSTs reaching 3–5 °C above the 1982–2016 reference period4. -

Trout Fishing in Hillsdale July, 2012 Guy Winig, Member Hillsdale Conservation Advisory Council
Roe Jan Brown-released to fight another day. Trout Fishing in Hillsdale July, 2012 Guy Winig, member Hillsdale Conservation Advisory Council Trout are our canaries in a coalmine. They are an excellent indicator of stream health, requiring clean, cold, well-oxygenated water to thrive. Without very high quality surface waters, trout just cannot survive. Hillsdale is fortunate to have several miles of trout streams including the Roeliff Jansen Kill, Taconic Creek, Agawamuck Creek and Green River with numerous smaller and unnamed tributaries contributing to them. Our streams and their residents endure many perils including natural flood and drought cycles as well as man made impacts like pollution, invasive species, barriers and over –fishing. Erosion adds silt that smothers aquatic food sources and gravel spawning beds. Introduced Didymo algae (a.k.a. ‘Rock Snot’) now found in the Green River, also chokes the streambed. Loss of vegetation exposes water to heating and eliminates important breeding, feeding and cover areas for trout. Readers may recall the massive fish kill that resulted from the release of manure into the Roe Jan in the 90’s or tales of unscrupulous ‘sportsmen’ using small explosives and chlorine to kill all the fish in a segment of stream. Maybe the most harmful and insidious are non-point sources of pollution such as chemicals, fertilizers and heated runoff from parking lots, roads, roofs, crop fields and lawns. In 2009 and 2010, members of the community with help from the Columbia-Greene chapter of Trout Unlimited and the N.Y.S.D.E.C. Trees for Tributaries program planted several hundred small trees and shrubs on the banks and flood plain of the Roe Jan Kill through the Roe Jan Park in an effort to stabilize banks, introduce vegetative cover and provide shade to maintain cool stream temperatures. -

New and Emerging Technologies for Sustainable Fisheries: a Comprehensive Landscape Analysis
Photo by Pablo Sanchez Quiza New and Emerging Technologies for Sustainable Fisheries: A Comprehensive Landscape Analysis Environmental Defense Fund | Oceans Technology Solutions | April 2021 New and Emerging Technologies for Sustainable Fisheries: A Comprehensive Landscape Analysis Authors: Christopher Cusack, Omisha Manglani, Shems Jud, Katie Westfall and Rod Fujita Environmental Defense Fund Nicole Sarto and Poppy Brittingham Nicole Sarto Consulting Huff McGonigal Fathom Consulting To contact the authors please submit a message through: edf.org/oceans/smart-boats edf.org | 2 Contents List of Acronyms ...................................................................................................................................................... 5 1. Introduction .............................................................................................................................................................7 2. Transformative Technologies......................................................................................................................... 10 2.1 Sensors ........................................................................................................................................................... 10 2.2 Satellite remote sensing ...........................................................................................................................12 2.3 Data Collection Platforms ...................................................................................................................... -

Marine Fish Conservation Global Evidence for the Effects of Selected Interventions
Marine Fish Conservation Global evidence for the effects of selected interventions Natasha Taylor, Leo J. Clarke, Khatija Alliji, Chris Barrett, Rosslyn McIntyre, Rebecca0 K. Smith & William J. Sutherland CONSERVATION EVIDENCE SERIES SYNOPSES Marine Fish Conservation Global evidence for the effects of selected interventions Natasha Taylor, Leo J. Clarke, Khatija Alliji, Chris Barrett, Rosslyn McIntyre, Rebecca K. Smith and William J. Sutherland Conservation Evidence Series Synopses 1 Copyright © 2021 William J. Sutherland This work is licensed under a Creative Commons Attribution 4.0 International license (CC BY 4.0). This license allows you to share, copy, distribute and transmit the work; to adapt the work and to make commercial use of the work providing attribution is made to the authors (but not in any way that suggests that they endorse you or your use of the work). Attribution should include the following information: Taylor, N., Clarke, L.J., Alliji, K., Barrett, C., McIntyre, R., Smith, R.K., and Sutherland, W.J. (2021) Marine Fish Conservation: Global Evidence for the Effects of Selected Interventions. Synopses of Conservation Evidence Series. University of Cambridge, Cambridge, UK. Further details about CC BY licenses are available at https://creativecommons.org/licenses/by/4.0/ Cover image: Circling fish in the waters of the Halmahera Sea (Pacific Ocean) off the Raja Ampat Islands, Indonesia, by Leslie Burkhalter. Digital material and resources associated with this synopsis are available at https://www.conservationevidence.com/ -

Ecosystem Services Generated by Fish Populations
AR-211 Ecological Economics 29 (1999) 253 –268 ANALYSIS Ecosystem services generated by fish populations Cecilia M. Holmlund *, Monica Hammer Natural Resources Management, Department of Systems Ecology, Stockholm University, S-106 91, Stockholm, Sweden Abstract In this paper, we review the role of fish populations in generating ecosystem services based on documented ecological functions and human demands of fish. The ongoing overexploitation of global fish resources concerns our societies, not only in terms of decreasing fish populations important for consumption and recreational activities. Rather, a number of ecosystem services generated by fish populations are also at risk, with consequences for biodiversity, ecosystem functioning, and ultimately human welfare. Examples are provided from marine and freshwater ecosystems, in various parts of the world, and include all life-stages of fish. Ecosystem services are here defined as fundamental services for maintaining ecosystem functioning and resilience, or demand-derived services based on human values. To secure the generation of ecosystem services from fish populations, management approaches need to address the fact that fish are embedded in ecosystems and that substitutions for declining populations and habitat losses, such as fish stocking and nature reserves, rarely replace losses of all services. © 1999 Elsevier Science B.V. All rights reserved. Keywords: Ecosystem services; Fish populations; Fisheries management; Biodiversity 1. Introduction 15 000 are marine and nearly 10 000 are freshwa ter (Nelson, 1994). Global capture fisheries har Fish constitute one of the major protein sources vested 101 million tonnes of fish including 27 for humans around the world. There are to date million tonnes of bycatch in 1995, and 11 million some 25 000 different known fish species of which tonnes were produced in aquaculture the same year (FAO, 1997). -

Localized Plankton Blooms and Jubilees on the Gulf Coast Gordon Gunter Gulf Coast Research Laboratory
Gulf Research Reports Volume 6 | Issue 3 January 1979 Localized Plankton Blooms and Jubilees on the Gulf Coast Gordon Gunter Gulf Coast Research Laboratory Charles H. Lyles Gulf Coast Research Laboratory DOI: 10.18785/grr.0603.12 Follow this and additional works at: http://aquila.usm.edu/gcr Part of the Marine Biology Commons Recommended Citation Gunter, G. and C. H. Lyles. 1979. Localized Plankton Blooms and Jubilees on the Gulf Coast. Gulf Research Reports 6 (3): 297-299. Retrieved from http://aquila.usm.edu/gcr/vol6/iss3/12 This Short Communication is brought to you for free and open access by The Aquila Digital Community. It has been accepted for inclusion in Gulf and Caribbean Research by an authorized editor of The Aquila Digital Community. For more information, please contact [email protected]. Gulf Research Reports, Vol. 6,NO. 3,291-299, 1919. LOCALIZED PLANKTON BLOOMS AND JUBILEES ON THE GULF COAST GORDON GUNTER AND CHARLES H. LYLES Gulf Coast Research Laboratory, Ocean Springs, Mississippi 39564 and Gulf States Marine Fisheries Commission, Ocean Springs, Mississippi 39564 ABSTRACT The writers describe various small types of plankton blooms such as those occurring in boat slips, the head of a large bayou and a strip type bloom of Chaeroceras on the Gulf beach. Oyster kills from “poison water” draining off of marshes are said to be caused by plankton bloom. Small “jubilees” are said to be caused by localized blooms and one of these is described as it occurred. In November 1938, Dr. Margaretha Brongersma-Sanders of billions of Chaetoceras sp., a common open sea diatom. -

Seagrass Recovery Following Marine Heat Wave Influences Sediment Carbon Stocks
W&M ScholarWorks VIMS Articles Virginia Institute of Marine Science 1-2021 Seagrass Recovery Following Marine Heat Wave Influences Sediment Carbon Stocks Lillian R. Aoki Karen J. McGlathery Patricia L. Wiberg Matthew P. J. Oreska Amelie C. Berger See next page for additional authors Follow this and additional works at: https://scholarworks.wm.edu/vimsarticles Part of the Marine Biology Commons Recommended Citation Aoki, Lillian R.; McGlathery, Karen J.; Wiberg, Patricia L.; Oreska, Matthew P. J.; Berger, Amelie C.; Berg, Peter; and Orth, Robert J., Seagrass Recovery Following Marine Heat Wave Influences Sediment Carbon Stocks (2021). Frontiers in Marine Science, 7, 576784.. doi: 10.3389/fmars.2020.576784 This Article is brought to you for free and open access by the Virginia Institute of Marine Science at W&M ScholarWorks. It has been accepted for inclusion in VIMS Articles by an authorized administrator of W&M ScholarWorks. For more information, please contact [email protected]. Authors Lillian R. Aoki, Karen J. McGlathery, Patricia L. Wiberg, Matthew P. J. Oreska, Amelie C. Berger, Peter Berg, and Robert J. Orth This article is available at W&M ScholarWorks: https://scholarworks.wm.edu/vimsarticles/2036 fmars-07-576784 December 23, 2020 Time: 12:35 # 1 ORIGINAL RESEARCH published: 07 January 2021 doi: 10.3389/fmars.2020.576784 Seagrass Recovery Following Marine Heat Wave Influences Sediment Carbon Stocks Lillian R. Aoki1*†, Karen J. McGlathery1, Patricia L. Wiberg1, Matthew P. J. Oreska1, Amelie C. Berger1, Peter Berg1 and Robert J. Orth2 1 Department of Environmental Sciences, University of Virginia, Charlottesville, VA, United States, 2 Virginia Institute of Marine Science, William and Mary, Gloucester Point, VA, United States Worldwide, seagrass meadows accumulate significant stocks of organic carbon (C), known as “blue” carbon, which can remain buried for decades to centuries. -

Fish Passage Engineering Design Criteria 2019
FISH PASSAGE ENGINEERING DESIGN CRITERIA 2019 37.2’ U.S. Fish and Wildlife Service Northeast Region June 2019 Fish and Aquatic Conservation, Fish Passage Engineering Ecological Services, Conservation Planning Assistance United States Fish and Wildlife Service Region 5 FISH PASSAGE ENGINEERING DESIGN CRITERIA June 2019 This manual replaces all previous editions of the Fish Passage Engineering Design Criteria issued by the U.S. Fish and Wildlife Service Region 5 Suggested citation: USFWS (U.S. Fish and Wildlife Service). 2019. Fish Passage Engineering Design Criteria. USFWS, Northeast Region R5, Hadley, Massachusetts. USFWS R5 Fish Passage Engineering Design Criteria June 2019 USFWS R5 Fish Passage Engineering Design Criteria June 2019 Contents List of Figures ................................................................................................................................ ix List of Tables .................................................................................................................................. x List of Equations ............................................................................................................................ xi List of Appendices ........................................................................................................................ xii 1 Scope of this Document ....................................................................................................... 1-1 1.1 Role of the USFWS Region 5 Fish Passage Engineering ............................................ -

Patterns and Processes in the Evolutionary History of Parrotfishes
bs_bs_banner Biological Journal of the Linnean Society, 2012, ••, ••–••. With 5 figures Patterns and processes in the evolutionary history of parrotfishes (Family Labridae) JOHN. H. CHOAT1*, OYA. S. KLANTEN1†, LYNNE VAN HERWERDEN1, D. ROSS ROBERTSON2‡ and KENDALL D. CLEMENTS3 1School of Tropical and Marine Biology, James Cook University, Townsville, QLD, 4811, Australia 2Smithsonian Tropical Research Institute, Ancon, Balboa, Republic of Panama 3School of Biological Sciences, University of Auckland, Private Bag 92019, Auckland 1142, New Zealand Received 5 March 2012; revised 23 May 2012; accepted for publication 23 May 2012 Phylogenetic reconstruction of the evolutionary relationships among 61 of the 70 species of the parrotfish genera Chlorurus and Scarus (Family Labridae) based on mitochondrial and nuclear gene sequences retrieved 15 well-supported clades with mid Pliocene/Pleistocene diversification. Twenty-two reciprocally monophyletic sister- species pairs were identified: 64% were allopatric, and the remainder were sympatric. Age of divergence was similar for allopatric and sympatric species pairs. Sympatric sister pairs displayed greater divergence in morphol- ogy, ecology, and sexually dimorphic colour patterns than did allopatric pairs, suggesting that both genetic drift in allopatric species pairs and ecologically adaptive divergence between members of sympatric pairs have played a role in diversification. Basal species typically have small geographical ranges and are restricted to geographically and ecologically peripheral reef habitats. We found little evidence that a single dominant process has driven diversification, nor did we detect a pattern of discrete, sequential stages of diversification in relation to habitat, ecology, and reproductive biology. The evolution of Chlorurus and Scarus has been complex, involving a number of speciation processes. -

The Tale of a Surprisingly Cold Blob in the North Atlantic
VARIATIONSUS CLIVAR VARIATIONS CUS CLIVAR lim ity a bil te V cta ariability & Predi Spring 2016 • Vol. 14, No. 2 A Tale of Two Blobs The evolution and known atmospheric Editors: forcing mechanisms behind the 2013-2015 Kristan Uhlenbrock & Mike Patterson North Pacific warm anomalies From 2013 to 2015, the scientific 1 2 community and the media were Dillon J. Amaya Nicholas E. Bond , enthralled with two anomalous Arthur J. Miller1, and Michael J. DeFlorio3 sea surface temperature events, both getting the moniker 1Scripps Institution of Oceanography the “Blob,” although one was 2 warm and one was cold. These University of Washington 3 events occurred during a Jet Propulsion Laboratory, California Institute of Technology period of record-setting global mean surface temperatures. This edition focuses on the timing and extent, possible mechanisms, and impacts ear-to-year variations in the El Niño Southern Oscillation (ENSO) indices of these unusual ocean heat Ygenerate significant interest throughout the general public and the scientific anomalies, and what we might community due to the sometimes destructive nature of this climate mode. For expect in the future as climate example, so-called “Godzilla” ENSOs can generate billions of dollars in damages changes. from the US agricultural industry alone due to unanticipated flooding or drought The “Warm Blob” feature (Adams et al. 1999). However, in the winter of 2013/2014, North Pacific sea surface appeared in the North Pacific temperature (SST) anomalies exceeded three standard deviations above the mean during winter 2013 and was over a large region, shifting focus away from the tropics and onto the extratropics first identified by Nick Bond, as the associated atmospheric circulation patterns helped exacerbate the most University of Washington. -

FISHING NEWSLETTER 2020/2021 Table of Contents FWP Administrative Regions and Hatchery Locations
FISHING NEWSLETTER 2020/2021 Table of Contents FWP Administrative Regions and Hatchery Locations .........................................................................................3 Region 1 Reports: Northwest Montana ..........................................................................................................5 Region 2 Reports: West Central Montana .....................................................................................................17 Region 3 Reports: Southwest Montana ........................................................................................................34 Region 4 Reports: North Central Montana ...................................................................................................44 Region 5 Reports: South Central Montana ...................................................................................................65 Region 6 Reports: Northeast Montana ........................................................................................................73 Region 7 Reports: Southeast Montana .........................................................................................................86 Montana Fish Hatchery Reports: .......................................................................................................................92 Murray Springs Trout Hatchery ...................................................................................................................92 Washoe Park Trout Hatchery .......................................................................................................................93 -
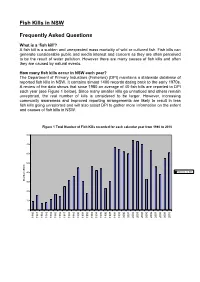
Fish Kills in NSW Frequently Asked Questions
Fish Kills in NSW Frequently Asked Questions What is a ‘fish kill’? A fish kill is a sudden and unexpected mass mortality of wild or cultured fish. Fish kills can generate considerable public and media interest and concern as they are often perceived to be the result of water pollution. However there are many causes of fish kills and often they are caused by natural events. How many fish kills occur in NSW each year? The Department of Primary Industries (Fisheries) (DPI) maintains a statewide database of reported fish kills in NSW. It contains almost 1400 records dating back to the early 1970s. A review of the data shows that since 1980 an average of 40 fish kills are reported to DPI each year (see Figure 1 below). Since many smaller kills go unnoticed and others remain unreported, the real number of kills is considered to be larger. However, increasing community awareness and improved reporting arrangements are likely to result in less fish kills going unreported and will also assist DPI to gather more information on the extent and causes of fish kills in NSW. Figure 1 Total Number of Fish Kills recorded for each calendar year from 1980 to 2010 80 70 60 50 40 Total no. of kills Number of kills 30 20 10 0 1980 1981 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 What species of fish are affected? Over 100 species of fish, including finfish, molluscs and crustaceans, have been reported in fish kills in NSW since records began.