Does Light Intensity Influence Song Output by Northern Mockingbirds? Christopher E
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Predation by Gray Catbird on Brown Thrasher Eggs
March 2004 Notes 101 PREDATION BY GRAY CATBIRD ON BROWN THRASHER EGGS JAMES W. RIVERS* AND BRETT K. SANDERCOCK Kansas Cooperative Fish and Wildlife Research Unit, Division of Biology, Kansas State University, Manhattan, KS 66506 (JWR) Division of Biology, Kansas State University, Manhattan, KS 66506 (BKS) Present address of JWR: Department of Ecology, Evolution, and Marine Biology, University of California, Santa Barbara, CA 93106 *Correspondent: [email protected] ABSTRACT The gray catbird (Dumetella carolinensis) has been documented visiting and breaking the eggs of arti®cial nests, but the implications of such observations are unclear because there is little cost in depredating an undefended nest. During the summer of 2001 at Konza Prairie Bio- logical Station, Kansas, we videotaped a gray catbird that broke and consumed at least 1 egg in a brown thrasher (Toxostoma rufum) nest. Our observation was consistent with egg predation because the catbird consumed the contents of the damaged egg after breaking it. The large difference in body mass suggests that a catbird (37 g) destroying eggs in a thrasher (69 g) nest might risk injury if caught in the act of predation and might explain why egg predation by catbirds has been poorly documented. Our observation indicated that the catbird should be considered as an egg predator of natural nests and that single-egg predation of songbird nests should not be attributed to egg removal by female brown-headed cowbirds (Molothrus ater) without additional evidence. RESUMEN El paÂjaro gato gris (Dumetella carolinensis) ha sido documentado visitando y rompien- do los huevos de nidos arti®ciales, pero las implicaciones de dichas observaciones no son claras porque hay poco costo por depredar un nido sin defensa. -

Catbird, Gray
Mockingbirds and Thrashers — Family Mimidae 449 Mockingbirds and Thrashers — Family Mimidae Gray Catbird Dumetella carolinensis Though the Gray Catbird breeds west almost to the coast of British Columbia, it is only a rare vagrant to California—the bulk of the population migrates east of the Rocky Mountains. But the species is on the increase: of 107 reports accepted by the California Bird Records Committee 1884–1999, one third were in just the last four years of this interval. Similarly, of the 20 records of the Gray Catbird in San Diego County, 10 have come since initiation of the field work for this atlas in 1997. Migration: Half of San Diego County’s known cat- Photo by Anthony Mercieca birds have been fall migrants, occurring as early as 24 September (1976, one at Point Loma, S7, K. van Vuren, Cabrillo National Monument, Point Loma 11–17 July 1988 Luther et al. 1979). Besides eight fall records from Point (B. and I. Mazin, Pyle and McCaskie 1992) certainly was. Loma, there is one from the Tijuana River valley 7–8 November 1964 (the only specimen, SDNHM 35095), Winter: Three wintering Gray Catbirds have been report- one from a boat 15 miles off Oceanside 26 October 1983 ed from San Diego County, from Balboa Park (R9) (M. W. Guest, Bevier 1990), and two from Paso Picacho 16 December 1972 (P. Unitt) and from Point Loma 7 Campground (M20) 29 October 1988 (D. W. Aguillard, November 1983–13 March 1984 (V. P. Johnson, Roberson Pyle and McCaskie 1992) and 17 November 2002 (T. 1986) and 31 October 1999–21 January 2000 (D. -

Belize), and Distribution in Yucatan
University of Neuchâtel, Switzerland Institut of Zoology Ecology of the Black Catbird, Melanoptila glabrirostris, at Shipstern Nature Reserve (Belize), and distribution in Yucatan. J.Laesser Annick Morgenthaler May 2003 Master thesis supervised by Prof. Claude Mermod and Dr. Louis-Félix Bersier CONTENTS INTRODUCTION 1. Aim and description of the study 2. Geographic setting 2.1. Yucatan peninsula 2.2. Belize 2.3. Shipstern Nature Reserve 2.3.1. History and previous studies 2.3.2. Climate 2.3.3. Geology and soils 2.3.4. Vegetation 2.3.5. Fauna 3. The Black Catbird 3.1. Taxonomy 3.2. Description 3.3. Breeding 3.4. Ecology and biology 3.5. Distribution and threats 3.6. Current protection measures FIRST PART: BIOLOGY, HABITAT AND DENSITY AT SHIPSTERN 4. Materials and methods 4.1. Census 4.1.1. Territory mapping 4.1.2. Transect point-count 4.2. Sizing and ringing 4.3. Nest survey (from hide) 5. Results 5.1. Biology 5.1.1. Morphometry 5.1.2. Nesting 5.1.3. Diet 5.1.4. Competition and predation 5.2. Habitat use and population density 5.2.1. Population density 5.2.2. Habitat use 5.2.3. Banded individuals monitoring 5.2.4. Distribution through the Reserve 6. Discussion 6.1. Biology 6.2. Habitat use and population density SECOND PART: DISTRIBUTION AND HABITATS THROUGHOUT THE RANGE 7. Materials and methods 7.1. Data collection 7.2. Visit to others sites 8. Results 8.1. Data compilation 8.2. Visited places 8.2.1. Corozalito (south of Shipstern lagoon) 8.2.2. -

Birds of the East Texas Baptist University Campus with Birds Observed Off-Campus During BIOL3400 Field Course
Birds of the East Texas Baptist University Campus with birds observed off-campus during BIOL3400 Field course Photo Credit: Talton Cooper Species Descriptions and Photos by students of BIOL3400 Edited by Troy A. Ladine Photo Credit: Kenneth Anding Links to Tables, Figures, and Species accounts for birds observed during May-term course or winter bird counts. Figure 1. Location of Environmental Studies Area Table. 1. Number of species and number of days observing birds during the field course from 2005 to 2016 and annual statistics. Table 2. Compilation of species observed during May 2005 - 2016 on campus and off-campus. Table 3. Number of days, by year, species have been observed on the campus of ETBU. Table 4. Number of days, by year, species have been observed during the off-campus trips. Table 5. Number of days, by year, species have been observed during a winter count of birds on the Environmental Studies Area of ETBU. Table 6. Species observed from 1 September to 1 October 2009 on the Environmental Studies Area of ETBU. Alphabetical Listing of Birds with authors of accounts and photographers . A Acadian Flycatcher B Anhinga B Belted Kingfisher Alder Flycatcher Bald Eagle Travis W. Sammons American Bittern Shane Kelehan Bewick's Wren Lynlea Hansen Rusty Collier Black Phoebe American Coot Leslie Fletcher Black-throated Blue Warbler Jordan Bartlett Jovana Nieto Jacob Stone American Crow Baltimore Oriole Black Vulture Zane Gruznina Pete Fitzsimmons Jeremy Alexander Darius Roberts George Plumlee Blair Brown Rachel Hastie Janae Wineland Brent Lewis American Goldfinch Barn Swallow Keely Schlabs Kathleen Santanello Katy Gifford Black-and-white Warbler Matthew Armendarez Jordan Brewer Sheridan A. -

Wildland Fire in Ecosystems: Effects of Fire on Fauna
United States Department of Agriculture Wildland Fire in Forest Service Rocky Mountain Ecosystems Research Station General Technical Report RMRS-GTR-42- volume 1 Effects of Fire on Fauna January 2000 Abstract _____________________________________ Smith, Jane Kapler, ed. 2000. Wildland fire in ecosystems: effects of fire on fauna. Gen. Tech. Rep. RMRS-GTR-42-vol. 1. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 83 p. Fires affect animals mainly through effects on their habitat. Fires often cause short-term increases in wildlife foods that contribute to increases in populations of some animals. These increases are moderated by the animals’ ability to thrive in the altered, often simplified, structure of the postfire environment. The extent of fire effects on animal communities generally depends on the extent of change in habitat structure and species composition caused by fire. Stand-replacement fires usually cause greater changes in the faunal communities of forests than in those of grasslands. Within forests, stand- replacement fires usually alter the animal community more dramatically than understory fires. Animal species are adapted to survive the pattern of fire frequency, season, size, severity, and uniformity that characterized their habitat in presettlement times. When fire frequency increases or decreases substantially or fire severity changes from presettlement patterns, habitat for many animal species declines. Keywords: fire effects, fire management, fire regime, habitat, succession, wildlife The volumes in “The Rainbow Series” will be published during the year 2000. To order, check the box or boxes below, fill in the address form, and send to the mailing address listed below. -

GRAY CATBIRDS by Mike Scully
GRAY CATBIRDS By Mike Scully (This article first appeared in the August 2010 SAAS newsletter.) It is interesting how different cultures can evolve quite different perceptions of the same bird. The Gray Catbird (Dumetella carolinensis), for example, is seemingly one of our more descriptively named species. Their common call does indeed bring to mind the mewing of a domestic cat, but only to those familiar with cats. Arthur C. Bent, in his epochal work, “Life Histories of North American Birds,” states that the Chippewa people of the American Midwest called this species “the bird that cries with grief.” Bent puzzled in print how the Chippewa could find the pleasant song of the catbird mournful. I believe they were referring not to the song, but to the distinctive call. It can come as a minor paradigm shift when one realizes that the plaintive call of this species does indeed possess a dirge-like quality. Gray Catbird perched on American Beautyberry. Photo by Lora Reynolds. Certainly the catbird was familiar to many of the original inhabitants of North America, much as it remains today; an attractive gray bird, slipping low between brush tangles, the frequency of its songs and calls belying any impression of secrecy. The Gray Catbird possesses two attributes that doubtless help it succeed in human-altered landscapes; it prefers dense edge and early successional habitats, and it possesses a faculty for recognizing and rejecting the eggs of the Brown-headed Cowbird. The present breeding range includes the majority of North America, but they are absent from the Far North, the Florida Peninsula, much of the Southwest, the Great Basin and our Pacific Coast. -

Pre-Lesson Plan
Pre-Lesson Plan Prior to taking part in the Winged Migration program at Tommy Thompson Park it is recommended that you complete the following lessons to familiarize your students with some of the birds they might see and some of the concepts they will learn during their field trip. The lessons can easily be integrated into your Science, Language Arts, Social Studies and Physical Education programs. Part 1: Amazing Birds As a class, read the provided “Wanted” posters. The posters depict a very small sampling of some of the amazing feats and features of birds. To complement these readings, display the following websites so that students can see some of these birds “up close.” Common Loon http://www.schollphoto.com/gallery/thumbnails.php?album=1 Black-Capped Chickadee http://sdakotabirds.com/species_photos/black_capped_chickadee.htm Ruby-Throated Hummingbird http://www.surfbirds.com/cgi-bin/gallery/search2.cgi?species=Ruby- throated%20Hummingbird Downy Woodpecker http://www.pbase.com/billko/downy_woodpecker Great Horned Owl www.owling.com/Great_Horned.htm When you visit Tommy Thompson Park, you may see chickadees, hummingbirds, and woodpeckers. These birds all breed in southern Ontario. However, you probably will not see a Great Horned Owl, because this specific bird is usually flying around at night. Below is a list of some other birds students might see when they visit Tommy Thompson Park. Have them chose one bird each and write a “Wanted” poster for it, focusing on a cool fact about that bird. Some web sites that will help them get started -

Volume 2E - Revised Baseline Ecological Risk Assessment Hudson River Pcbs Reassessment
PHASE 2 REPORT FURTHER SITE CHARACTERIZATION AND ANALYSIS VOLUME 2E - REVISED BASELINE ECOLOGICAL RISK ASSESSMENT HUDSON RIVER PCBS REASSESSMENT NOVEMBER 2000 For U.S. Environmental Protection Agency Region 2 and U.S. Army Corps of Engineers Kansas City District Book 2 of 2 Tables, Figures and Plates TAMS Consultants, Inc. Menzie-Cura & Associates, Inc. PHASE 2 REPORT FURTHER SITE CHARACTERIZATION AND ANALYSIS VOLUME 2E- REVISED BASELINE ECOLOGICAL RISK ASSESSMENT HUDSON RIVER PCBs REASSESSMENT RI/FS CONTENTS Volume 2E (Book 1 of 2) Page TABLE OF CONTENTS ........................................................ i LIST OF TABLES ........................................................... xiii LIST OF FIGURES ......................................................... xxv LIST OF PLATES .......................................................... xxvi EXECUTIVE SUMMARY ...................................................ES-1 1.0 INTRODUCTION .......................................................1 1.1 Purpose of Report .................................................1 1.2 Site History ......................................................2 1.2.1 Summary of PCB Sources to the Upper and Lower Hudson River ......4 1.2.2 Summary of Phase 2 Geochemical Analyses .......................5 1.2.3 Extent of Contamination in the Upper Hudson River ................5 1.2.3.1 PCBs in Sediment .....................................5 1.2.3.2 PCBs in the Water Column ..............................6 1.2.3.3 PCBs in Fish .........................................7 -

Gray Catbird, Northern Mockingbird and Brown Thrasher
Wildlife Note — 51 LDR0103 Gray Catbird, Northern Mockingbird and Brown Thrasher by Chuck Fergus Gray Catbird These three species are among the most vocal of our birds. All belong to Family Mimidae, the “mimic thrushes,” or “mimids,” and they often imitate the calls of other spe- cies, stringing these remembered vocalizations into long, variable songs. Family Mimidae has more than 30 spe- cies, which are found only in the New World, with most inhabiting the tropics. The mimids have long tails and short, rounded wings. The three species in the Northeast are solitary (living singly, in pairs and in family groups rather than in flocks), feed mainly on the ground and in shrubs, and generally eat insects in summer and fruits in winter. The sexes look alike. Adults are preyed upon by owls, hawks, foxes and house cats, and their nests may be raided by snakes, blue jays, crows, grackles, raccoons, opossums and squirrels. Gray Catbird (Dumetella carolinensis) — The gray cat- bird is eight to nine inches long, smaller and more slen- der than a robin, an overall dark gray with a black cap Beetles, ants, caterpillars, and chestnut around the vent. Individuals often jerk their grasshoppers, crickets and other insects tails — up, down, and in circles. The species is named are common foodstuffs. Catbirds often forage on the for its mewling call, although catbirds also deliver other ground, using their bills to flick aside leaves and twigs sounds. They migrate between breeding grounds in the while searching for insects. eastern two-thirds of North America and wintering ar- Although not as talkative as the northern mocking- eas in the coastal Southeast and Central America. -

Factors Affecting Feeding and Brooding of Gray Catbird Nestlings
FACTORS AFFECTING FEEDING AND BROODING OF GRAY CATBIRD NESTLINGS ELLEN J. JOHNSONAND LOUIS B. BEST Departmentof Animal Ecology,Iowa State University,Ames, Iowa 50011 USA ABSTRACT.--GrayCatbirds (Dumetellacarolinensis) were studied to documentpatterns of feeding and brooding nestlingsand to relate theseto nestlingage, brood size, time of day, time of season,and weather. Major factors affecting feeding were nestling age and brood size. The averagenumber of food deliveries.brood-•. h-• increasedlinearly with the log of nestlingage for both male and femaleparents. Also, volumeof food delivered/feedingtrip increasedlinearly with nestling age. As brood size increased,feeding frequencies/brood increasedbut food deliveries/nestlingdecreased. Brooding and shadingwere affectedmainly by nestlingage and ambient temperature.Time spent brooding decreasedsubstantially as nestlingsgrew older. Shading was important when the sun shone directly on the nest. Femalesspent significantlymore time brooding nestlingswhen rain increased.As ambient temperature increased, time spent shading increased and time spent brooding declined. Received7 May 1980, accepted27 April 1981. ONE of the greateststresses on birds during STUDY AREA AND METHODS the breeding seasonis caring for their nest- lings. Frequent feeding trips are required to This study was conductedin a shrubby and partly satisfythe food demandsof the rapidly devel- wooded pasture near Ames, Iowa during the springs and summers of 1977 and 1978. The area covered 16 oping young, and nestling body temperature ha of which approximately50% containedshrubs, must be maintained at a relatively constant primarily gooseberry(Ribes spp.) and multiflora rose level. Ratesof feeding and brooding by parent (Rosa multiflora), suitable as nesting sites for cat- birds can be related either to requirements of birds. A stream meandered through the study site. -

MD-Birds-2021-Envirothon-3Pp
3/23/2021 Maryland Envirothon: Class Aves KERRY WIXTED WILDLIFE AND HERITAGE SERVICE March 2021 1 Aves Overview •> 450 species in Maryland •Extirpated species include: ◦Bewick’s Wren ◦Greater Prairie Chicken ◦Red-cockaded Woodpecker American Woodcock (Scolopax minor) chick by Kerry Wixted Note: This guide is an overview of select species found in Maryland. The taxonomy and descriptions are based off Peterson Field Guide to Birds of Eastern and Central North America, 6th Ed. 2 Order: Anseriformes • Web-footed waterfowl • Family Anatidae • Ducks, geese & swans Male Wood Duck (Aix sponsa) Order: Anseriformes; Family Anatidae 3 1 3/23/2021 Cygnus- Swans Mute Swan Tundra Swan Trumpeter Swan (Cygnus olor) (Cygnus columbianus) (Cygnus buccinator) Invasive. Adult black-knobbed orange Winter resident. Adult bill black, usually Winter resident. Adult all black bill bill tilts down; immature is dingy with with small yellow basal spot; immature with straight ridge; more nasal calls is dingy in color with pinkish bill; makes than tundra swan pinkish bill; makes hissing sounds mellow-high pitched woo-ho, woo-woo noise Order: Anseriformes; Family Anatidae By Matthew Beziat CC by NC 2.0 By Jen Goellnitz CC by NC 2.0 4 Dabbling Ducks- feed by dabbling & upending Male Female , Maryland Biodiversity Project Biodiversity ,Maryland Hubick By Bill Bill By By Kerry Wixted Kerry By American Black Duck (Anas rubripes) Mallard (Anas platyrhynchos) Year-round resident. Dusky black duck with white wing linings Year-round resident. Adult males have green head w/ evident in flight; resembles a female mallard but has black white neck ring & females are mottled in color ; borders on secondary wing feathers; can hybridize with resembles an American Black Duck mallard but has mallards white borders on secondary wing feathers Order: Anseriformes; Family Anatidae 5 Diving Ducks- feed by diving; legs close to tail CC by NC NC 2.0CC by Beziat By Judy Gallagher CC by by 2.0CC Gallagher Judy By By Matthew Matthew By Canvasback (Aythya valisneria) Redhead (Aythya americana) Winter resident. -
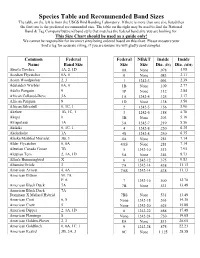
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American