Spatial Distribution of the Endangered Pacific Pocket Mouse (Perognathus Longimembrus Ssp
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
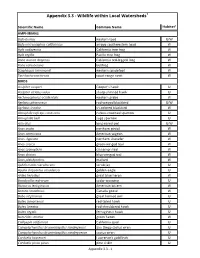
Appendix 3.3 - Wildlife Within Local Watersheds1
Appendix 3.3 - Wildlife within Local Watersheds1 2 Scientific Name Common Name Habitat AMPHIBIANS Bufo boreas western toad U/W Bufo microscaphus californicus arroyo southwestern toad W Hyla cadaverina California tree frog W Hyla regilla Pacific tree frog W Rana aurora draytonii California red-legged frog W Rana catesbeiana bullfrog W Scaphiopus hammondi western spadefoot W Taricha torosa torosa coast range newt W BIRDS Accipiter cooperi Cooper's hawk U Accipiter striatus velox sharp-shinned hawk U Aechmorphorus occidentalis western grebe W Agelaius phoeniceus red-winged blackbird U/W Agelaius tricolor tri-colored blackbird W Aimophila ruficeps canescens rufous-crowned sparrow U Aimophilia belli sage sparrow U Aiso otus long-eared owl U/W Anas acuta northern pintail W Anas americana American wigeon W Anas clypeata northern shoveler W Anas crecca green-winged teal W Anas cyanoptera cinnamon teal W Anas discors blue-winged teal W Anas platrhynchos mallard W Aphelocoma coerulescens scrub jay U Aquila chrysaetos canadensis golden eagle U Ardea herodius great blue heron W Bombycilla cedrorum cedar waxwing U Botaurus lentiginosus American bittern W Branta canadensis Canada goose W Bubo virginianus great horned owl U Buteo jamaicensis red-tailed hawk U Buteo lineatus red-shouldered hawk U Buteo regalis ferruginous hawk U Butorides striatus green heron W Callipepla californica California quail U Campylorhynchus brunneicapillus sandiegensis San Diego cactus wren U Campylorhynchus brunneicapillus sandiegoense cactus wren U Carduelis lawrencei Lawrence's -

Field Guide of Common Ants
J U L Y / A U G 2 0 0 3 N P M A L I B R A R Y U P D A T E Insert this update into the NPMA Pest Management Library, which can be Updatepurchased from the Resource Center. phone: 703-573-8330 fax: 703-573-4116 Common Ants: A Pull-Out Guide for Use in the Field This Library Update is designed to of acrobat ants. Workers are 1/16” to 1/8” assist technicians in identification and in length and are normally brown to control of ants while servicing accounts. black in color. The pedicel, or front joint This update is not designed for of the abdomen has two nodes or instruction in basic ant biology, segments. Looking down on the ant nomenclature of the anatomy of the ant, under hand lens magnification, one pair or to be used as a key. For more detailed of spines is found on the thorax. Various information on those topics, refer to the species of acrobat ants are found Field Guide or other technical materials. throughout the United States. In the field, a great aid to identification Acrobat ants are named for their is the use of a hand lens. Many of these tendency to raise their abdomens in the ants are small and positive identification air when disturbed. As the abdomen is is not easy without a hand lens. This heart shaped and is frequently black and article focuses on common, non-wood- shiny, the acrobat motion is readily destroying ants, including: acrobat, observable. -

Body Size, Not Phylogenetic Relationship Or Residency, Drives Interspecific Dominance in a Little Pocket Mouse Community
Animal Behaviour 137 (2018) 197e204 Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav Body size, not phylogenetic relationship or residency, drives interspecific dominance in a little pocket mouse community * Rachel Y. Chock a, , Debra M. Shier a, b, Gregory F. Grether a a Department of Ecology & Evolutionary Biology, University of California, Los Angeles, CA, U.S.A. b Recovery Ecology, San Diego Zoo Institute for Conservation Research, Escondido, CA, U.S.A. article info The role of interspecific aggression in structuring ecological communities can be important to consider Article history: when reintroducing endangered species to areas of their historic range that are occupied by competitors. Received 6 September 2017 We sought to determine which species is the most serious interference competitor of the endangered Initial acceptance 6 November 2017 Pacific pocket mouse, Perognathus longimembris pacificus, and more generally, whether interspecific Final acceptance 1 December 2017 aggression in rodents is predicted by body size, residency status or phylogenetic relatedness. We carried out simulated territory intrusion experiments between P. longimembris and four sympatric species of MS. number: A17-00719 rodents (Chaetodipus fallax, Dipodomys simulans, Peromyscus maniculatus, Reithrodontomys megalotis)ina field enclosure in southern California sage scrub habitat. We found that body size asymmetries strongly Keywords: predicted dominance, regardless of phylogenetic relatedness or the residency status of the individuals. aggression The largest species, D. simulans, was the most dominant while the smallest species, R. megalotis, was the dominance least dominant to P. longimembris. Furthermore, P. longimembris actively avoided encounters with all interference competition Perognathus longimembris species, except R. -

Taxonomy and Distribution of the Argentine Ant, Linepithema Humile (Hymenoptera: Formicidae)
SYSTEMATICS Taxonomy and Distribution of the Argentine Ant, Linepithema humile (Hymenoptera: Formicidae) ALEXANDER L. WILD Department of Entomology, University of California at Davis, Davis, CA 95616 Ann. Entomol. Soc. Am. 97(6): 1204Ð1215 (2004) ABSTRACT The taxonomy of an invasive pest species, the Argentine ant, is reviewed. Linepithema humile (Mayr) 1868 is conÞrmed as the valid name for the Argentine ant. Morphological variation and species boundaries of L. humile are examined, with emphasis on populations from the antÕs native range in southern South America. Diagnoses and illustrations are provided for male, queen, and worker castes. Collection records of L. humile in South America support the idea of a native distribution closely associated with major waterways in lowland areas of the Parana´ River drainage, with recent intro- ductions into parts of Argentina, Brazil, Chile, Colombia, Ecuador, and Peru. KEY WORDS Argentine ant, Linepithema humile, taxonomy, invasive species THE ARGENTINE ANT, Linepithema humile (Mayr) 1868, MCSN, MCZC, MHNG, MZSP, NHMB, NHMW, and is among the worldÕs most successful invasive species. USNM; see below for explanation of abbreviations). This native South American insect has become a cos- Taxonomic confusion over L. humile extends be- mopolitan pest, particularly in the Mediterranean cli- yond museum collections. At least one important mates of North America, Chile, South Africa, Austra- study, seeking to explain Argentine ant population lia, and southern Europe (Suarez et al. 2001). regulation in the native range through phorid para- Argentine ants have been implicated in the decline of sitism (Orr and Seike 1998), initially targeted the native arthropod (Cole et al. 1992) and vertebrate wrong Linepithema species (Orr et al. -

The Natural World That I Seek out in the Desert Regions of Baja California
The natural world that I seek out in the desert regions of Baja California and southern California provides me with scientific adventure, excitement towards botany, respect for nature, and overall feelings of peace and purpose. Jon P. Rebman, Ph.D. has been the Mary and Dallas Clark Endowed Chair/Curator of Botany at the San Diego Natural History Museum (SDNHM) since 1996. He has a Ph.D. in Botany (plant taxonomy), M.S. in Biology (floristics) and B.S. in Biology. Dr. Rebman is a plant taxonomist and conducts extensive floristic research in Baja California and in San Diego and Imperial Counties. He has over 15 years of experience in the floristics of San Diego and Imperial Counties and 21 years experience studying the plants of the Baja California peninsula. He leads various field classes and botanical expeditions each year and is actively naming new plant species from our region. His primary research interests have centered on the systematics of the Cactus family in Baja California, especially the genera Cylindropuntia (chollas) and Opuntia (prickly-pears). However, Dr. Rebman also does a lot of general floristic research and he co- published the most recent edition of the Checklist of the Vascular Plants of San Diego County. He has over 22 years of field experience with surveying and documenting plants including rare and endangered species. As a field botanist, he is a very active collector of scientific specimens with his personal collections numbering over 22,500. Since 1996, he has been providing plant specimen identification/verification for various biological consulting companies on contracts dealing with plant inventory projects and environmental assessments throughout southern California. -

Sauromalus Hispidus
ARTÍCULOS CIENTÍFICOS Cerdá-Ardura & Langarica-Andonegui 2018 - Sauromalus hispidus in Rasa Island- p 17-28 ON THE PRESENCE OF THE SPINY CHUCKWALLA SAUROMALUS HISPIDUS (STEJNEGER, 1891) IN RASA ISLAND, MEXICO PRESENCIA DEL CHACHORÓN ESPINOSO SAUROMALUS HISPIDUS (STEJNEGER, 1891) EN LA ISLA DE RASA, MÉXICO Adrián Cerdá-Ardura1* and Esther Langarica-Andonegui2 1Lindblad Expeditions/National Geographic. 2Facultad de Ciencias, Uiversidad Nacional Autónoma de México, CDMX, México. *Correspondence author: [email protected] Abstract.— In 2006 and 2013 two different individuals of the Spiny Chuckwalla (Sauromalus hispidus) were found on the small, flat, volcanic and isolated Rasa Island, located in the Midriff Region of the Gulf of California, Mexico. This species had never been recorded from Rasa Island prior to 2006. A new field study in 2014 revealed the presence of a single female chuckwalla inhabiting the Tapete Verde Valley, in the south-central part of the island, occupying a territory no bigger than 10000 m2. A scat analysis shows that the only food consumed by the animal is the Alkali Weed (Cressa truxilliensis) that forms patches of carpets in its habitat. The individual is in precarious condition, as it seems to starve on a seasonal basis, especially during El Niño cycles; also, it is missing fingers and toes, which appear to be intentional markings by amputation. We conclude that the two individuals were introduced to the island intentionally by humans. Keywords.— Chuckwalla, Gulf of California, Rasa Island. Resumen.— En 2006 y 2013 se encontraron dos individuos diferentes del cachorón de roca o chuckwalla espinoso (Sauromalus hispidus) en la pequeña, plana, volcánica y aislada isla Rasa, localizada en la Región de las Grandes Islas, en el Golfo de California, México. -

Draft Environmental Assessment of Marine Geophysical Surveys by the R/V Marcus G. Langseth for the Southern California Collaborative Offshore Geophysical Survey
DRAFT ENVIRONMENTAL ASSESSMENT OF MARINE GEOPHYSICAL SURVEYS BY THE R/V MARCUS G. LANGSETH FOR THE SOUTHERN CALIFORNIA COLLABORATIVE OFFSHORE GEOPHYSICAL SURVEY Submitted to: National Science Foundation Division of Ocean Sciences 4201 Wilson Blvd., Suite 725 Arlington, VA 22230 Submitted by: Scripps Institution of Oceanography, UCSD 8675 Discovery Way La Jolla, CA 92023 Contact: Professor Neal Driscoll 858.822.5026; [email protected] Prepared by: Padre Associates, Inc. 5290 Overpass Road, Suite 217 Goleta, CA 93113 June 2012 Southern California Collaborative Offshore Geophysical Survey (SCCOGS) Environmental Assessment TABLE OF CONTENTS 1.0 PURPOSE AND NEED ................................................................................................... 1 2.0 ALTERNATIVES INCLUDING PROPOSED ACTION ...................................................... 6 2.1 PROPOSED ACTION ......................................................................................... 6 2.2 PROJECT LOCATION ........................................................................................ 6 2.3 PROJECT ACTIVITIES ....................................................................................... 6 2.3.1 Mobilization and Demobilization .............................................................. 9 2.3.2 Offshore Survey Operations .................................................................... 9 2.3.2.1 Survey Vessel Specifications ..................................................... 10 2.3.2.2 Air Gun Description ................................................................... -

LOS ANGELES POCKET MOUSE Perognathus Longimembris Brevinasus
Terrestrial Mammal Species of Special Concern in California, Bolster, B.C., Ed., 1998 111 Los Angeles pocket mouse, Perognathus longimembris brevinasus Philip V. Brylski Description: This is a small heteromyid rodent, averaging about 113 mm TL with weight from 8 to 11 g. The Los Angeles pocket mouse can be potentially confused only with juveniles of the sympatric California pocket mouse (Chaetodipus californicus), from which it can be distinguished by the absence of spiny hairs in the dorsal pelage and the absence of a distinct crest on the tail. Pelage is buff above and white below. Many of the dorsal hairs are black-tipped, giving the pelage a "salt and pepper" appearance, similar to but lighter than that of the Pacific pocket mouse (P. l. pacificus). Like all silky pocket mice, there is usually a small white spot at the anterior base of the ear, and an indistinct larger buff spot behind the ear, the plantar surface of the hindfeet are naked or lightly haired, and the lateral hairs of the hind toes project anteriorly and laterally, resulting in a "fringed-toed" effect, which may enhance locomotor efficiency on sandy substrates. Taxonomic Remarks: This is one of eight subspecies of the little pocket mouse (P. longimembris) in California (Hall 1981). P. l. brevinasus was first described by Osgood (1900) as a race of the Panamint pocket mouse (P. panamintinus). Both brevinasus and panamintinus were arranged as subspecies of P. longimembris by Huey (1928). An important taxonomic character of brevinasus is its short rostrum, a character also shared by pacificus. P. -

The Case of the Argentine Ant in Vineyards of Northern Argentina
insects Article Growing Industries, Growing Invasions? The Case of the Argentine Ant in Vineyards of Northern Argentina Maria Schulze-Sylvester 1,* ID , José A. Corronca 1 ID and Carolina I. Paris 2 1 FCN-IEBI (Instituto para el Estudio de la Biodiversidad de Invertebrados), CONICET CCT-Salta, Facultad de Ciencias Naturales, Universidad Nacional de Salta, Av. Bolivia 5150, CP 4400 Salta, Argentina; [email protected] 2 Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Buenos Aires, 1428 Buenos Aires, Argentina; [email protected] * Correspondence: [email protected]; Tel.: +54-387-425-5472 Received: 10 December 2017; Accepted: 23 January 2018; Published: 29 January 2018 Abstract: The invasive Argentine ant causes ecological and economic damage worldwide. In 2011, this species was reported in vineyards of Cafayate, a wine-producing town in the Andes, Argentina. While the local xeric climate is unsuitable for Argentine ants, populations could establish in association with vineyards where human activity and irrigation facilitate propagule introduction and survival. In 2013–2014, we combined extensive sampling of the area using ant-baits with monitoring of the change in land use and vineyard cultivated area over the past 15 years. Our results revealed that the species has thus far remained confined to a relatively isolated small area, owing to an effective barrier of dry shrublands surrounding the infested vineyards; yet the recent expansion of vineyard acreage in this region will soon connect this encapsulated area with the rest of the valley. When this happens, vulnerable ecosystems and the main local industry will be put at risk. -

Taxonomy and Distribution of Opuntia and Related Plants
Taxonomy and Distribution of Opuntia and Related Genera Raul Puente Desert Botanical Garden Donald Pinkava Arizona State University Subfamily Opuntioideae Ca. 350 spp. 13-18 genera Very wide distribution (Canada to Patagonia) Morphological consistency Glochids Bony arils Generic Boundaries Britton and Rose, 1919 Anderson, 2001 Hunt, 2006 -- Seven genera -- 15 genera --18 genera Austrocylindropuntia Austrocylindropuntia Grusonia Brasiliopuntia Brasiliopuntia Maihuenia Consolea Consolea Nopalea Cumulopuntia Cumulopuntia Opuntia Cylindropuntia Cylindropuntia Pereskiopsis Grusonia Grusonia Pterocactus Maihueniopsis Corynopuntia Tacinga Miqueliopuntia Micropuntia Opuntia Maihueniopsis Nopalea Miqueliopuntia Pereskiopsis Opuntia Pterocactus Nopalea Quiabentia Pereskiopsis Tacinga Pterocactus Tephrocactus Quiabentia Tunilla Tacinga Tephrocactus Tunilla Classification: Family: Cactaceae Subfamily: Maihuenioideae Pereskioideae Cactoideae Opuntioideae Wallace, 2002 Opuntia Griffith, P. 2002 Nopalea nrITS Consolea Tacinga Brasiliopuntia Tunilla Miqueliopuntia Cylindropuntia Grusonia Opuntioideae Grusonia pulchella Pereskiopsis Austrocylindropuntia Quiabentia 95 Cumulopuntia Tephrocactus Pterocactus Maihueniopsis Cactoideae Maihuenioideae Pereskia aculeata Pereskiodeae Pereskia grandiflora Talinum Portulacaceae Origin and Dispersal Andean Region (Wallace and Dickie, 2002) Cylindropuntia Cylindropuntia tesajo Cylindropuntia thurberi (Engelmann) F. M. Knuth Cylindropuntia cholla (Weber) F. M. Knuth Potential overlapping areas between the Opuntia -

Recovery Research for the Endangered Pacific Pocket Mouse: an Overview of Collaborative Studies1
Recovery Research for the Endangered Pacific Pocket Mouse: An Overview of Collaborative Studies1 Wayne D. Spencer2 Abstract The critically endangered Pacific pocket mouse (Perognathus longimembris pacificus), feared extinct for over 20 years, was “rediscovered” in 1993 and is now documented at four sites in Orange and San Diego Counties, California. Only one of these sites is considered large enough to be potentially self-sustaining without active intervention. In 1998, I gathered a team of biologists to initiate several research tasks in support of recovery planning for the species. The PPM Studies Team quickly determined that species recovery would require active trans- locations or reintroductions to establish new populations, but that we knew too little about the biology of P. l. pacificus and the availability of translocation receiver sites to design such a program. Recovery research from 1998 to 2000 therefore focused on (1) a systematic search for potential translocation receiver sites; (2) laboratory and field studies on non-listed, surro- gate subspecies (P. l. longimembris and P. l. bangsi) to gain biological insights and perfect study methods; (3) studies on the historic and extant genetic diversity of P. l. pacificus; and (4) experimental habitat manipulations to increase P. l. pacificus populations. Using existing geographic information system (GIS) data, we identified sites throughout the historic range that might have appropriate soils and vegetation to support translocated P. l. pacificus. Re- connaissance surveys of habitat value were completed in all large areas of potential habitat identified by the model. Those sites having the highest habitat potential are being studied with more detailed and quantitative field analyses. -

US EPA-Pesticides; Disodium Octaborate Tetrahydrate
EFFICACY REVIEW DATE: IN 4-30-01 OUT 6-21-01 FILE OR REG. NO. 69529-1 PETITION OR EXP. PERMIT NO. DATE DIV. RECEIVED April 19, 2001 DATE OF SUBMISSION April 18, 2001 DATE SUBMISSION ACCEPTED TYPE PRODUCT(S): (I,)D, H, F, N, R, S DATA ACCESSION NO(S). 453568-01;D274488;S596027;Case#046539;AC:301 PRODUCT MGR. NO. 03-Layne/Quarles PRODUCT NAME(S) Pestbor COMPANY NAME Quality Borate Company SUBMISSION PURPOSE Provide performance data in reprint supporting claims for Argentine ant, pharaoh ant and Tap- inoma melanocephalum for formulated products. CHEMICAL & FORMULATION Disodium octaborate tetrahydrate 98% (30 lbs./cu.ft. bulk density manufacturing concentrate) CONCLUSIONS & RECOMMENDATIONS The data presented in EPA Accession (MRID) Number 453568-01, having been obtained from the reprint art- icle titled “Laboratory Evaluation of a Boric Acid Liquid Bait on Colonies of Tapinoma melanocephalum, Argentine Ants and Pharaoh Ants (Hymenoptera: Formicidae)S, which meets the requirements of § 11(b)(1)-(7) on p. 268 and the standard of § 95-11(c)(3)(b) on pp. 270-1 of the Product Performance Guidelines, are adequate to sup- port the registration of the subject product, the sole use for this formulation being for the manufacturing of end use insecticidal products, more specifically baits. The cited data (to be contin'd) is far more than is necessary to establish usefulness of this form- ulation for the making of insecticidal baits. Results with the 3 species included in the testing were as follows: Tapinoma melanocephalum workers were reduced by 97%