2 Biomimetic Microsystems for Blood and Lymphatic Vascular Research
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vocabulario De Morfoloxía, Anatomía E Citoloxía Veterinaria
Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) Servizo de Normalización Lingüística Universidade de Santiago de Compostela COLECCIÓN VOCABULARIOS TEMÁTICOS N.º 4 SERVIZO DE NORMALIZACIÓN LINGÜÍSTICA Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) 2008 UNIVERSIDADE DE SANTIAGO DE COMPOSTELA VOCABULARIO de morfoloxía, anatomía e citoloxía veterinaria : (galego-español- inglés) / coordinador Xusto A. Rodríguez Río, Servizo de Normalización Lingüística ; autores Matilde Lombardero Fernández ... [et al.]. – Santiago de Compostela : Universidade de Santiago de Compostela, Servizo de Publicacións e Intercambio Científico, 2008. – 369 p. ; 21 cm. – (Vocabularios temáticos ; 4). - D.L. C 2458-2008. – ISBN 978-84-9887-018-3 1.Medicina �������������������������������������������������������������������������veterinaria-Diccionarios�������������������������������������������������. 2.Galego (Lingua)-Glosarios, vocabularios, etc. políglotas. I.Lombardero Fernández, Matilde. II.Rodríguez Rio, Xusto A. coord. III. Universidade de Santiago de Compostela. Servizo de Normalización Lingüística, coord. IV.Universidade de Santiago de Compostela. Servizo de Publicacións e Intercambio Científico, ed. V.Serie. 591.4(038)=699=60=20 Coordinador Xusto A. Rodríguez Río (Área de Terminoloxía. Servizo de Normalización Lingüística. Universidade de Santiago de Compostela) Autoras/res Matilde Lombardero Fernández (doutora en Veterinaria e profesora do Departamento de Anatomía e Produción Animal. -

27 Lymph Vascular System
Lymph Vascular System The lymph vascular system consists of endothelial-lined tubes that recover intercellular (tissue) fluid not picked up by the blood vascular system and returns it to the blood. The fluid (lymph) carried by the lymphatics is a blood filtrate formed as fluid crosses the blood capillaries into the tissues. Unlike the blood vascular system, lymph flow is unidirectional - from tissues to the union of the lymphatic and blood vascular systems at the base of the neck. The lymphatic vascular system begins in the tissues as blindly ending capillaries that drain into larger collecting vessels and then into two main lymphatic trunks. Lymph nodes occur along the course of the vessels and filter the lymph. Lymphatics are present in most tissues but are absent from bone marrow, the central nervous system, coats of the eye, internal ear, and fetal placenta. Lymph Capillaries Lymph capillaries are thin-walled, blind tubes that branch and anastomose freely to form a rich network in organs and tissues. They are wider and more irregular than blood capillaries. The wall of a lymph capillary consists only of a thin continuous endothelium and a discontinuous basal lamina that is present only in patches or may even be absent. Adjacent endothelial cells may overlap, but junctional complexes are few and clefts occur between the cells. Externally, the endothelium is surrounded by a small amount of collagenous connective tissue. Fine filaments run perpendicularly from the collagen bundles and attach to the outer surfaces of the endothelium as anchoring filaments that maintain the patency of the vessel. Collecting Lymph Vessels Collecting lymph vessels differ from lymph capillaries in size and the thickness of their walls. -

Structure, Function, and Molecular Control of the Skin Lymphatic System
CORE Metadata, citation and similar papers at core.ac.uk Provided by Elsevier - Publisher Connector Structure, Function, and Molecular Control of the Skin Lymphatic System Mihaela Skobe and Michael Detmar Cutaneous Biology Research Center, Department of Dermatology, Massachusetts General Hospital and Harvard Medical School, Charlestown, Massachusetts, U.S.A. The mechanisms of angiogenesis have been studied phatic vasculature and the differences between blood extensively over the past years. The focus, however, and lymphatic vessels. Special attention has been has been almost exclusively on blood vessels, whereas given to the methods employed in research of the little effort has been directed toward understanding lymphatic system. Finally, we describe molecular lymphangiogenesis and the role of lymphatic vessels mechanisms involved in the regulation of lymphan- in physiology and pathology. The lymphatic system, giogenesis. Vascular endothelial growth factor and acting in concert with the blood vascular system, is vascular endothelial growth factor-C, expressed by of fundamental importance in maintaining tissue distinct skin cell populations, play an important role homeostasis, and disorders of the lymphatic system in the molecular control of skin angiogenesis and are common, often resulting in chronic, disabling lymphangiogenesis. Key words: lymphatic vessels/lymph- conditions. This overview summarizes the most angiogenesis/skin/VEGF-C. Journal of Investigative important aspects of the structure and function of Dermatology Symposium Proceedings 5:14±19, 2000 the lymphatic system with emphasis on the skin lym- ur understanding of the biology of the lymphatic covered'' (Bartels, 1909). The ancient Greeks observed structures system is well illustrated by the word lymphatic containing colorless ¯uid (Hippocrates spoke of ``white blood'') but itself; the derivation of the latin word lymphaticus their function was not understood and the signi®cance of the signi®es ``distracted and confused'' (Witte et al, ®nding was not recognized. -

Lymphatic Tissue Engineering and Regeneration Laura Alderfer1, Alicia Wei1 and Donny Hanjaya-Putra1,2,3,4,5,6*
Alderfer et al. Journal of Biological Engineering (2018) 12:32 https://doi.org/10.1186/s13036-018-0122-7 REVIEW Open Access Lymphatic Tissue Engineering and Regeneration Laura Alderfer1, Alicia Wei1 and Donny Hanjaya-Putra1,2,3,4,5,6* Abstract The lymphatic system is a major circulatory system within the body, responsible for the transport of interstitial fluid, waste products, immune cells, and proteins. Compared to other physiological systems, the molecular mechanisms and underlying disease pathology largely remain to be understood which has hindered advancements in therapeutic options for lymphatic disorders. Dysfunction of the lymphatic system is associated with a wide range of disease phenotypes and has also been speculated as a route to rescue healthy phenotypes in areas including cardiovascular disease, metabolic syndrome, and neurological conditions. This review will discuss lymphatic system functions and structure, cell sources for regenerating lymphatic vessels, current approaches for engineering lymphatic vessels, and specific therapeutic areas that would benefit from advances in lymphatic tissue engineering and regeneration. Keywords: Lymphangiogenesis, Tissue Engineering, Disease Modeling, Wound Healing, Lymphedema, Stem Cells, Biomaterials, Interstitial Fluid, Regeneration I. Introduction to the Lymphatic System and its role Interstitial fluid (IF) is a plasma filtrate that is generated Function by transcapillary filtration and is governed by Starling The lymphatic system is nearly ubiquitous in the human forces, the net difference between hydrostatic and body, present in all tissues except the epidermis, cartil- osmotic pressures, at the microcirculatory level [9]. In age, eye lens, cornea, retina, and bone marrow [1, 2]. order to maintain fluid homeostasis, lymph formation in The main functions of the lymphatic system include the initial lymphatic vessels must be balanced by the net fluid homeostasis and interstitial fluid drainage, immune flux of plasma being filtered out [4]. -

A Role of Inflammation and Immunity in Essential Hypertension—Modeled and Analyzed Using Petri Nets
International Journal of Molecular Sciences Article A Role of Inflammation and Immunity in Essential Hypertension—Modeled and Analyzed Using Petri Nets Dorota Formanowicz 1 , Agnieszka Rybarczyk 2,3 , Marcin Radom 2,3 and Piotr Formanowicz 2,3,* 1 Department of Clinical Biochemistry and Laboratory Medicine, Poznan University of Medical Sciences, 60-806 Poznan, Poland; [email protected] 2 Institute of Computing Science, Poznan University of Technology, 60-965 Poznan, Poland; [email protected] (A.R.); [email protected] (M.R.) 3 Institute of Bioorganic Chemistry, Polish Academy of Sciences, 61-704 Poznan, Poland * Correspondence: [email protected] Received: 15 April 2020; Accepted: 5 May 2020; Published: 9 May 2020 Abstract: Recent studies have shown that the innate and adaptive immune system, together with low-grade inflammation, may play an important role in essential hypertension. In this work, to verify the importance of selected factors for the development of essential hypertension, we created a Petri net-based model and analyzed it. The analysis was based mainly on t-invariants, knockouts of selected fragments of the net and its simulations. The blockade of the renin-angiotensin (RAA) system revealed that the most significant effect on the emergence of essential hypertension has RAA activation. This blockade affects: (1) the formation of angiotensin II, (2) inflammatory process (by influencing C-reactive protein (CRP)), (3) the initiation of blood coagulation, (4) bradykinin generation via the kallikrein-kinin system, (5) activation of lymphocytes in hypertension, (6) the participation of TNF alpha in the activation of the acute phase response, and (7) activation of NADPH oxidase—a key enzyme of oxidative stress. -
Lymphatic System Urls
Lymphatic System URLs Human Anatomy & Physiology 16 http://www.howstuffworks.com/immune-system.htm http://www.thebody.com/step/immune.html http://www.emc.maricopa.edu/faculty/farabee/BIOBK/ BioBookIMMUN.html & http://www.cayuga-cc.edu/about/facultypages/greer/ r http://www.acm.uiuc.edu/sigbio/project/updated- lymphatic/lymph1.html http://www.pblsh.com/Healthworks/lymphart.html Karen Webb Smith Unit Fou Introduction A. The lymphatic system is closely associated with the cardiovascular system and is comprised of a network of vessels that circulate body fluids. B. Lymphatic vessels transport excess fluid away from interstitial spaces between cells in most tissues & return it to the bloodstream. C. Lymphatic vessels called lacteals (located in the in the lining of the smallsmall intestine) absorb fats resulting from digestion, & then transport fats to the circulatory system. D. The organs of the lymphatic system help defend Lymphatic vessels against disease. transporting fluid from interstitial spaces to the bloodstream Lymphatic Pathways A. Lymphatic pathways start as lymphatic capillaries that merge to form larger vessels that empty into the circulatory system. (This is key to understanding this chapter.) B. Lymphatic Capillaries *are microscopic, close-ended tubes that extend into interstitial spaces forming networks that parallel the networks of the blood capillaries *walls consist of single layer squamous epithelial cells which enables interstitial fluid to enter the lymphatic capillaries *lymph – the fluid inside a lymph capillary C. Lymphatic Vessels *walls of lymphatic vessels are thinner than walls of veins * have semilunar valves to prevent backflow of lymph *lymph nodes – specialized lymph organs that are composed of a mass of lymphoid tissue located along the course of a lymphatic vessel D. -

Human and Nonhuman Primate Meninges Harbor Lymphatic Vessels
SHORT REPORT Human and nonhuman primate meninges harbor lymphatic vessels that can be visualized noninvasively by MRI Martina Absinta1†, Seung-Kwon Ha1†, Govind Nair1, Pascal Sati1, Nicholas J Luciano1, Maryknoll Palisoc2, Antoine Louveau3, Kareem A Zaghloul4, Stefania Pittaluga2, Jonathan Kipnis3, Daniel S Reich1* 1Translational Neuroradiology Section, National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, United States; 2Hematopathology Section, Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bethesda, United States; 3Center for Brain Immunology and Glia, Department of Neuroscience, School of Medicine, University of Virginia, Charlottesville, United States; 4Surgical Neurology Branch, National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, United States Abstract Here, we report the existence of meningeal lymphatic vessels in human and nonhuman primates (common marmoset monkeys) and the feasibility of noninvasively imaging and mapping them in vivo with high-resolution, clinical MRI. On T2-FLAIR and T1-weighted black-blood imaging, lymphatic vessels enhance with gadobutrol, a gadolinium-based contrast agent with high propensity to extravasate across a permeable capillary endothelial barrier, but not with gadofosveset, a blood-pool contrast agent. The topography of these vessels, running alongside dural venous sinuses, recapitulates the meningeal lymphatic system of rodents. In primates, *For correspondence: meningeal -

New Brain Lymphatic Vessels Drain Old Concepts
EBioMedicine 2 (2015) 776–777 Contents lists available at ScienceDirect EBioMedicine journal homepage: www.ebiomedicine.com In Focus New Brain Lymphatic Vessels Drain Old Concepts Lasse Dissing-Olesen, Soyon Hong, Beth Stevens⁎ Department of Neurology, F.M. Kirby Neurobiology Center, Boston Children's Hospital, Harvard Medical School, Boston, MA 02115, USA The brain has traditionally been considered an immune privileged vessels above the dcLN completely abolished drainage of dye to these organ, in part due to the lack of evidence for lymphatic vasculature lymph nodes and additionally, increased the diameter of the meningeal (Galea et al., 2007). Two centuries ago, the existence of lymphatic ves- lymphatic vessels, indicating that these vessels drain to the dcLN. The sels on the surface of the human brain was proposed but it has since Alitalo lab further demonstrated that dye injected into the brain paren- been widely dismissed (Lukic et al., 2003). Therefore, while T cells chyma was absorbed by lymphatic vessels and preferentially drained leave all other organs via the lymphatic system to reach nearby lymph into the ipsilateral dcLN from the base of the brain. Surprisingly, Kipnis' nodes, the prevailing view has been that infiltrated T cells exit the lab excluded the best-known route of drainage into the dcLN, via the brain via venous blood circulation, circumventing the lymph nodes nasal cavity's cribriform plate and the nasal mucosa (Kida et al., 1995), (Ransohoff and Engelhardt, 2012). The recent evidence for the existence as they failed -

Extraperitoneal Laparoscopic Resection for Retroperitoneal Lymphatic Cysts
Wang et al. BMC Urology (2017) 17:101 DOI 10.1186/s12894-017-0288-1 RESEARCHARTICLE Open Access Extraperitoneal laparoscopic resection for retroperitoneal lymphatic cysts: initial experience Yichun Wang1†, Chen Chen1†, Chuanjie Zhang1†, Chao Qin2* and Ninghong Song2* Abstract Background: To assess the safety and efficacy of laparoscopic retroperitoneal resection for retroperitoneal lymphatic cysts. Methods: A retrospective analysis was conducted based on clinical data from eight patients with hydronephrosis caused by retroperitoneal lymphatic cysts. All patients underwent laparoscopic retroperitoneal lymphatic cyst resection and received postoperative follow-up. A follow-up ultrasound was performed postoperatively every 6–12 months to evaluate the recovery of the hydronephrosis. Results: All operations were successful, and their postoperative pathological results revealed lymphatic cyst walls. The operation time ranged from 43 to 88 min (mean: 62 min), with a blood loss of 20 to 130 mL (mean: 76 mL), and the length of hospital stay was 3 to 6 days (mean: 4.5 days). Within the follow-up of 12 to 36 months (mean: 28.5 months), great relief was detected in all eight cases, and no recurrence was found. Moreover, complications such as renal pedicle or renal pelvis injury were not observed. Conclusions: Laparoscopic retroperitoneal lymphatic cyst resection is an effective treatment for retroperitoneal lymphatic cysts and has the advantages of being minimally invasive, producing less intraoperative blood loss and leading to a quick recovery. This treatment thus deserves further studies. Keywords: Hydronephrosis, Lymphatic cyst, Laparoscope Background pain, obstructive uropathy, lower lymphoedema, bowel Lymphatic cysts are a rare lymphatic-vessel-generated obstruction and venous thrombosis [7–10]. -

Índice De Denominacións Españolas
VOCABULARIO Índice de denominacións españolas 255 VOCABULARIO 256 VOCABULARIO agente tensioactivo pulmonar, 2441 A agranulocito, 32 abaxial, 3 agujero aórtico, 1317 abertura pupilar, 6 agujero de la vena cava, 1178 abierto de atrás, 4 agujero dental inferior, 1179 abierto de delante, 5 agujero magno, 1182 ablación, 1717 agujero mandibular, 1179 abomaso, 7 agujero mentoniano, 1180 acetábulo, 10 agujero obturado, 1181 ácido biliar, 11 agujero occipital, 1182 ácido desoxirribonucleico, 12 agujero oval, 1183 ácido desoxirribonucleico agujero sacro, 1184 nucleosómico, 28 agujero vertebral, 1185 ácido nucleico, 13 aire, 1560 ácido ribonucleico, 14 ala, 1 ácido ribonucleico mensajero, 167 ala de la nariz, 2 ácido ribonucleico ribosómico, 168 alantoamnios, 33 acino hepático, 15 alantoides, 34 acorne, 16 albardado, 35 acostarse, 850 albugínea, 2574 acromático, 17 aldosterona, 36 acromatina, 18 almohadilla, 38 acromion, 19 almohadilla carpiana, 39 acrosoma, 20 almohadilla córnea, 40 ACTH, 1335 almohadilla dental, 41 actina, 21 almohadilla dentaria, 41 actina F, 22 almohadilla digital, 42 actina G, 23 almohadilla metacarpiana, 43 actitud, 24 almohadilla metatarsiana, 44 acueducto cerebral, 25 almohadilla tarsiana, 45 acueducto de Silvio, 25 alocórtex, 46 acueducto mesencefálico, 25 alto de cola, 2260 adamantoblasto, 59 altura a la punta de la espalda, 56 adenohipófisis, 26 altura anterior de la espalda, 56 ADH, 1336 altura del esternón, 47 adipocito, 27 altura del pecho, 48 ADN, 12 altura del tórax, 48 ADN nucleosómico, 28 alunarado, 49 ADNn, 28 -

Lateral Peritumoral Lymphatic Vessel Invasion Can Predict Lymph Node Metastasis in Esophageal Squamous Cell Carcinoma
Modern Pathology (2007) 20, 694–700 & 2007 USCAP, Inc All rights reserved 0893-3952/07 $30.00 www.modernpathology.org Lateral peritumoral lymphatic vessel invasion can predict lymph node metastasis in esophageal squamous cell carcinoma Daisuke Mori1, Fumio Yamasaki1, Masami Shibaki1 and Osamu Tokunaga2 1Division of Pathology, Saga Prefectural Hospital Kouseikan, Saga, Japan and 2Faculty of Medicine, Department of Pathology and Biodefense, Saga University, Saga, Japan Lymph node metastasis is an important prognostic factor in many types of cancer. We investigated the clinical significance of lymphangiogenesis and lymphatic vessel invasion in esophageal squamous cell carcinoma. We evaluated lymphatic vessel density and lymphatic vessel invasion in the intratumoral, peritumoral and normal compartments using D2-40 immunostaining. In addition, the peritumoral compartment was divided into the lateral peritumoral compartment and the non-lateral peritumoral compartment. The lymphatic vessel density was higher in the peritumoral and intratumoral compartments than in the normal compartment. However, the lymphatic vessel density did not correlate with any pathological parameters including lymph node metastasis. Intratumoral and peritumoral lymph vessels were small and collapsed while normal lymphatic vessels and lymphatic vessels with lymphatic vessel invasion were dilated and large. The presence of lymphatic vessel invasion, in the lateral peritumoral compartment but nowhere else, significantly correlated with lymph node metastasis. These results -
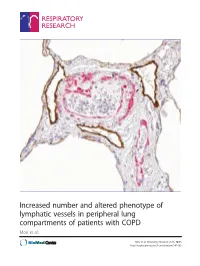
Increased Number and Altered Phenotype of Lymphatic Vessels in Peripheral Lung Compartments of Patients with COPD Mori Et Al
Increased number and altered phenotype of lymphatic vessels in peripheral lung compartments of patients with COPD Mori et al. Mori et al. Respiratory Research 2013, 14:65 http://respiratory-research.com/content/14/1/65 Mori et al. Respiratory Research 2013, 14:65 http://respiratory-research.com/content/14/1/65 RESEARCH Open Access Increased number and altered phenotype of lymphatic vessels in peripheral lung compartments of patients with COPD Michiko Mori1, Cecilia K Andersson2, Gerard J Graham3, Claes-Göran Löfdahl2 and Jonas S Erjefält1,2* Abstract Background: De novo lymphatic vessel formation has recently been observed in lungs of patients with moderate chronic obstructive pulmonary disease (COPD). However, the distribution of lymphatic vessel changes among the anatomical compartments of diseased lungs is unknown. Furthermore, information regarding the nature of lymphatic vessel alterations across different stages of COPD is missing. This study performs a detailed morphometric characterization of lymphatic vessels in major peripheral lung compartments of patients with different severities of COPD and investigates the lymphatic expression of molecules involved in immune cell trafficking. Methods: Peripheral lung resection samples obtained from patients with mild (GOLD stage I), moderate-severe (GOLD stage II-III), and very severe (GOLD stage IV) COPD were investigated for podoplanin-immunopositive lymphatic vessels in distinct peripheral lung compartments: bronchioles, pulmonary blood vessels and alveolar walls. Control subjects with normal lung function were divided into never smokers and smokers. Lymphatics were analysed by multiple morphological parameters, as well as for their expression of CCL21 and the chemokine scavenger receptor D6. Results: The number of lymphatics increased by 133% in the alveolar parenchyma in patients with advanced COPD compared with never-smoking controls (p < 0.05).