Cell Size Predicts Morphological Complexity in the Brains of Frogs and Salamanders GERHARD ROTH*T, JENS BLANKE*, and DAVID B
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca
Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca John F. Lamoreux, Meghan W. McKnight, and Rodolfo Cabrera Hernandez Occasional Paper of the IUCN Species Survival Commission No. 53 Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca John F. Lamoreux, Meghan W. McKnight, and Rodolfo Cabrera Hernandez Occasional Paper of the IUCN Species Survival Commission No. 53 The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN or other participating organizations. Published by: IUCN, Gland, Switzerland Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Citation: Lamoreux, J. F., McKnight, M. W., and R. Cabrera Hernandez (2015). Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca. Gland, Switzerland: IUCN. xxiv + 320pp. ISBN: 978-2-8317-1717-3 DOI: 10.2305/IUCN.CH.2015.SSC-OP.53.en Cover photographs: Totontepec landscape; new Plectrohyla species, Ixalotriton niger, Concepción Pápalo, Thorius minutissimus, Craugastor pozo (panels, left to right) Back cover photograph: Collecting in Chamula, Chiapas Photo credits: The cover photographs were taken by the authors under grant agreements with the two main project funders: NGS and CEPF. -
Comparative Osteology and Evolution of the Lungless Salamanders, Family Plethodontidae David B
COMPARATIVE OSTEOLOGY AND EVOLUTION OF THE LUNGLESS SALAMANDERS, FAMILY PLETHODONTIDAE DAVID B. WAKE1 ABSTRACT: Lungless salamanders of the family Plethodontidae comprise the largest and most diverse group of tailed amphibians. An evolutionary morphological approach has been employed to elucidate evolutionary rela tionships, patterns and trends within the family. Comparative osteology has been emphasized and skeletons of all twenty-three genera and three-fourths of the one hundred eighty-three species have been studied. A detailed osteological analysis includes consideration of the evolution of each element as well as the functional unit of which it is a part. Functional and developmental aspects are stressed. A new classification is suggested, based on osteological and other char acters. The subfamily Desmognathinae includes the genera Desmognathus, Leurognathus, and Phaeognathus. Members of the subfamily Plethodontinae are placed in three tribes. The tribe Hemidactyliini includes the genera Gyri nophilus, Pseudotriton, Stereochilus, Eurycea, Typhlomolge, and Hemidac tylium. The genera Plethodon, Aneides, and Ensatina comprise the tribe Pleth odontini. The highly diversified tribe Bolitoglossini includes three super genera. The supergenera Hydromantes and Batrachoseps include the nominal genera only. The supergenus Bolitoglossa includes Bolitoglossa, Oedipina, Pseudoeurycea, Chiropterotriton, Parvimolge, Lineatriton, and Thorius. Manculus is considered to be congeneric with Eurycea, and Magnadig ita is congeneric with Bolitoglossa. Two species are assigned to Typhlomolge, which is recognized as a genus distinct from Eurycea. No. new information is available concerning Haptoglossa. Recognition of a family Desmognathidae is rejected. All genera are defined and suprageneric groupings are defined and char acterized. Range maps are presented for all genera. Relationships of all genera are discussed. -

Rediscovery at the Type Locality and Detection of a New Population 1José L
Offcial journal website: Amphibian & Reptile Conservation amphibian-reptile-conservation.org 13(2) [General Section]: 126–132 (e193). Thorius narismagnus (Amphibia: Plethodontidae): rediscovery at the type locality and detection of a new population 1José L. Aguilar-López, 2,*Paulina García-Bañuelos, 3Eduardo Pineda, and 4Sean M. Rovito 1,2,3Red de Biología y Conservación de Vertebrados, Instituto de Ecología, A.C., Carretera antigua a Coatepec 351, El Haya, Xalapa, Veracruz, MEXICO 4Unidad de Genómica Avanzada (Langebio), Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, km 9.6 Libramiento Norte Carretera Irapuato-León, Irapuato, Guanajuato CP 36824, MEXICO Abstract.—Of the 42 Critically Endangered species of plethodontid salamanders that occur in Mexico, thirteen have not been reported in more than ten years. Given the lack of reports since 1976, the minute plethodontid salamander Thorius narismagnus is widely considered as missing. However, this report describes the rediscovery of this minute salamander at the type locality (Volcán San Martín), as well as a new locality on Volcán Santa Marta, 28 km southeast of its previously known distribution, both in the Los Tuxtlas region of Veracruz, Mexico. The localities where T. narismagnus has been found are mature forests in a community reserve on Volcán San Martín and a private reserve on Volcán Santa Marta. The presence of maxillary teeth, generally absent in Thorius, are reported here in some T. narismagnus females. Two efforts which may contribute to the conservation of Thorius narismagnus are the preservation of the cloud forests where this species persists, as well as the determination of the presence and possible effect of chytrid fungus in these populations. -

Pseudoeurycea Naucampatepetl. the Cofre De Perote Salamander Is Endemic to the Sierra Madre Oriental of Eastern Mexico. This
Pseudoeurycea naucampatepetl. The Cofre de Perote salamander is endemic to the Sierra Madre Oriental of eastern Mexico. This relatively large salamander (reported to attain a total length of 150 mm) is recorded only from, “a narrow ridge extending east from Cofre de Perote and terminating [on] a small peak (Cerro Volcancillo) at the type locality,” in central Veracruz, at elevations from 2,500 to 3,000 m (Amphibian Species of the World website). Pseudoeurycea naucampatepetl has been assigned to the P. bellii complex of the P. bellii group (Raffaëlli 2007) and is considered most closely related to P. gigantea, a species endemic to the La specimens and has not been seen for 20 years, despite thorough surveys in 2003 and 2004 (EDGE; www.edgeofexistence.org), and thus it might be extinct. The habitat at the type locality (pine-oak forest with abundant bunch grass) lies within Lower Montane Wet Forest (Wilson and Johnson 2010; IUCN Red List website [accessed 21 April 2013]). The known specimens were “found beneath the surface of roadside banks” (www.edgeofexistence.org) along the road to Las Lajas Microwave Station, 15 kilometers (by road) south of Highway 140 from Las Vigas, Veracruz (Amphibian Species of the World website). This species is terrestrial and presumed to reproduce by direct development. Pseudoeurycea naucampatepetl is placed as number 89 in the top 100 Evolutionarily Distinct and Globally Endangered amphib- ians (EDGE; www.edgeofexistence.org). We calculated this animal’s EVS as 17, which is in the middle of the high vulnerability category (see text for explanation), and its IUCN status has been assessed as Critically Endangered. -
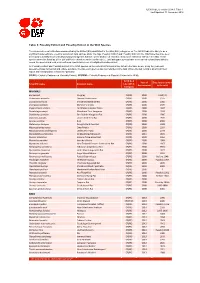
Table 9: Possibly Extinct and Possibly Extinct in the Wild Species
IUCN Red List version 2014.3: Table 9 Last Updated: 13 November 2014 Table 9: Possibly Extinct and Possibly Extinct in the Wild Species The number of recent extinctions documented by the Extinct (EX) and Extinct in the Wild (EW) categories on The IUCN Red List is likely to be a significant underestimate, even for well-known taxa such as birds. The tags 'Possibly Extinct' and 'Possibly Extinct in the Wild' have therefore been developed to identify those Critically Endangered species that are, on the balance of evidence, likely to be extinct (or extinct in the wild). These species cannot be listed as EX or EW until their extinction can be confirmed (i.e., until adequate surveys have been carried out and have failed to record the species and local or unconfirmed reports have been investigated and discounted). All 'Possibly Extinct' and 'Possibly Extinct in the Wild' species on the current IUCN Red List are listed in the table below, along the year each assessment was carried out and, where available, the date each species was last recorded in the wild. Where the last record is an unconfirmed report, last recorded date is noted as "possibly". CR(PE) - Critically Endangered (Possibly Extinct), CR(PEW) - Critically Endangered (Possibly Extinct in the Wild), IUCN Red Year of Date last recorded Scientific name Common name List (2014) Assessment in the wild Category MAMMALS Bos sauveli Kouprey CR(PE) 2008 1969/70 Crateromys australis Dinagat Crateromys CR(PE) 2008 1975 Crocidura trichura Christmas Island Shrew CR(PE) 2008 1985 Crocidura wimmeri -

Results of the Global Amphibian Assessment for Bolivia
Results of the Global Amphibian Assessment for Bolivia Diversity Number of Rank in Latin World Rank2 Percentage of Species in Bolivia America and the the World’s Caribbean1 Diversity All Amphibians 201 7 15 3.5 % Frogs & Toads 197 7 14 3.9 % Salamanders 1 13 68 0.2 % Caecilians 3 11 20 1.8 % 1 Out of 44 countries and territories. 2 Out of 192 countries and territories. Threatened Species (Threatened species are in one of the categories in italics.) IUCN Categories Number of Species in IUCN Categories Number of Species in Bolivia Bolivia Extinct 0 Near Threatened 6 Critically Endangered 5 Least Concern 161 Endangered 6 Data Deficient 13 Vulnerable 10 Number and percent of Bolivian species that are threatened: 21 (10%) Extinct Species: none Species that are Critically Endangered and possibly extinct (species that are missing but for which sufficient information is lacking to declare them as extinct): Frogs: Gastrotheca lauzuricae, Hyla chlorostea, Eleutherodactylus zongoensis Role of Protected Areas: Of the 21 threatened species in Bolivia, 76% occur in at least one protected area. Local Experts: Steffen Reichle, The Nature Conservancy, TEL (591-3) 348 0766, 348 0767 Claudia Cortez, Colección Boliviana de Fauna, TEL (591-3) 272 1152 For More Information: Dr. Bruce Young, NatureServe, Costa Rica, TEL (506) 645-6231 Global Amphibian Assessment website: www.globalamphibians.org A detailed analysis of the results for the Americas: www.natureserve.org Results of the Global Amphibian Assessment for Bolivia Map of Amphibian Diversity Source: Global Amphibian Assessment Map of the Distribution of Threatened Species Source: Global Amphibian Assessment Results of the Global Amphibian Assessment for Brazil Diversity Number of Rank in Latin World Rank2 Percentage of Species in Brazil America and the the World’s Caribbean1 Diversity All Amphibians 731 1 1 12.7 % Frogs & Toads 704 1 1 13.9 % Salamanders 1 13 68 0.2 % Caecilians 26 2 2 15.5 % 1 Out of 44 countries and territories. -

Functional Morphology and Evolution of Tail Autotomy in Salamanders
Functional Morphology and Evolution of Tail Autotomy in Salamanders DAVID B. WAKE AND IAN G. DRESNER Department of Anatomy, The University of Chicago, Chicago, lllinois ABSTRACT Basal tail constriction occurs in about two-thirds of the species of plethodontid salamanders. The constriction, which marks the site of tail autotomy, is a result of a reduction in length and diameter of the first caudal segment. Gross and microscopic anatomical studies reveal that many structural specializations are associated with basal constriction, and these are considered in detail. Areas of weak- ness in the skin at the posterior end of the first caudal segment, at the attachment of the musculature to the intermyotomal septum at the anterior end of the same segment, and between the last caudosacral and first caudal vertebrae precisely define the route of tail breakage. During autotomy the entire tail is shed, and a cylinder of skin one segment long closes over the wound at the end of the body. It is suggested that specializations described in this paper have evolved jndepend- ently in three different groups of salamanders. Experiments and field observations reveal that, contrary to expectations, frequency of tail breakage is less in species with apparent provisions for tail autotomy than in less specialized species. The tail is a very important, highly functional organ in salamanders and it is suggested that selection has been for behavioral and structural adaptations for control of tail loss, rather than for tail loss per se. Tail loss and subsequent regeneration is sent facts concerning anatomical details a well documented phenomenon among of the basal tail region, a functional inter- the lower vertebrates. -

I Online Supplementary Data – Sexual Size Dimorphism in Salamanders
Online Supplementary data – Sexual size dimorphism in salamanders Supplementary data S1. Species data used in this study and references list. Males Females SSD Significant test Ref Species n SVL±SD n SVL±SD Andrias davidianus 2 532.5 8 383.0 -0.280 12 Cryptobranchus alleganiensis 53 277.4±5.2 52 300.9±3.4 0.084 Yes 61 Batrachuperus karlschmidti 10 80.0 10 84.8 0.060 26 Batrachuperus londongensis 20 98.6 10 96.7 -0.019 12 Batrachuperus pinchonii 5 69.6 5 74.6 0.070 26 Batrachuperus taibaiensis 11 92.9±12.1 9 102.1±7.1 0.099 Yes 27 Batrachuperus tibetanus 10 94.5 10 92.8 -0.017 12 Batrachuperus yenyuadensis 10 82.8 10 74.8 -0.096 26 Hynobius abei 24 57.8±2.1 34 55.0±1.2 -0.048 Yes 92 Hynobius amakusaensis 22 75.4±4.8 12 76.5±3.6 0.014 No 93 Hynobius arisanensis 72 54.3±4.8 40 55.2±4.8 0.016 No 94 Hynobius boulengeri 37 83.0±5.4 15 91.5±3.8 0.102 Yes 95 Hynobius formosanus 15 53.0±4.4 8 52.4±3.9 -0.011 No 94 Hynobius fuca 4 50.9±2.8 3 52.8±2.0 0.037 No 94 Hynobius glacialis 12 63.1±4.7 11 58.9±5.2 -0.066 No 94 Hynobius hidamontanus 39 47.7±1.0 15 51.3±1.2 0.075 Yes 96 Hynobius katoi 12 58.4±3.3 10 62.7±1.6 0.073 Yes 97 Hynobius kimurae 20 63.0±1.5 15 72.7±2.0 0.153 Yes 98 Hynobius leechii 70 61.6±4.5 18 66.5±5.9 0.079 Yes 99 Hynobius lichenatus 37 58.5±1.9 2 53.8 -0.080 100 Hynobius maoershanensis 4 86.1 2 80.1 -0.069 101 Hynobius naevius 72.1 76.7 0.063 102 Hynobius nebulosus 14 48.3±2.9 12 50.4±2.1 0.043 Yes 96 Hynobius osumiensis 9 68.4±3.1 15 70.2±3.0 0.026 No 103 Hynobius quelpaertensis 41 52.5±3.8 4 61.3±4.1 0.167 Yes 104 Hynobius -

Hanken and Wake 1998 Copeia.Pdf
Copeia, 1998(2), pp. 312-345 Biology of Tiny Animals: Systematics of the Minute Salamanders (Thorius: Plethodontidae) from Veracruz and Puebla, Mexico, with Descriptions of Five New Species JAMES HANKEN AND DAVID B. WAKE Minute plethodontid salamanders, genus Thorius, are far more diverse taxonom- ically than has been recognized previously. Populations of these salamanders from the Mexican states of Veracruz and Puebla are assigned to 10 species, five of which are described as new. Combinations of morphological and allozymic characters are used to sort the species and to make initial assessments of relationships. Valid ex- isting names include Thorius pennatulus, T. troglodytes, T. dubitus, and T. schmidti. Thorius narismagnus, from the Sierra de Los Tuxtlas, which previously was consid- ered to be a disjunct subspecies of T. pennatulus, is elevated to species rank. Thorius maxillabrochusis treated as a subjective junior synonym of the sympatric T. schmidti. New taxa include Thorius lunaris, T. magnipes, T. minydemus, T. munificus, and T. spilogaster. All 10 species can be distinguished by morphological characters, but the distinctiveness of the taxa is bolstered by allozymic characters and by extensive sympatry. As many as three, and possibly four, species occur in sympatry, with some evidence of segregation by microhabitat (arboreal vs terrestrial). Adult body sizes span the range known for the genus, from very small in T. pennatulus (maturing at < 16 mm standard length) to large in T. lunaris (adults reaching > 31 mm). Collec- tively these species display a wide elevational distribution, from less than 1000 m (T. pennatulus, T. narismagnus) to more than 3000 m (T lunaris, T. -

Rampant Tooth Loss Across 200 Million Years of Frog Evolution
bioRxiv preprint doi: https://doi.org/10.1101/2021.02.04.429809; this version posted February 6, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. 1 Rampant tooth loss across 200 million years of frog evolution 2 3 4 Daniel J. Paluh1,2, Karina Riddell1, Catherine M. Early1,3, Maggie M. Hantak1, Gregory F.M. 5 Jongsma1,2, Rachel M. Keeffe1,2, Fernanda Magalhães Silva1,4, Stuart V. Nielsen1, María Camila 6 Vallejo-Pareja1,2, Edward L. Stanley1, David C. Blackburn1 7 8 1Department of Natural History, Florida Museum of Natural History, University of Florida, 9 Gainesville, Florida USA 32611 10 2Department of Biology, University of Florida, Gainesville, Florida USA 32611 11 3Biology Department, Science Museum of Minnesota, Saint Paul, Minnesota USA 55102 12 4Programa de Pós Graduação em Zoologia, Universidade Federal do Pará/Museu Paraense 13 Emilio Goeldi, Belém, Pará Brazil 14 15 *Corresponding author: Daniel J. Paluh, [email protected], +1 814-602-3764 16 17 Key words: Anura; teeth; edentulism; toothlessness; trait lability; comparative methods 1 bioRxiv preprint doi: https://doi.org/10.1101/2021.02.04.429809; this version posted February 6, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. -

Proceedings of the United States National Museum
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued i^?fv vl vJ^^S ^V '^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 95 Washington : 1945 No. 3185 SUMMARY OF THE COLLECTIONS OF AMPHIBIANS MADE IN MEXICO UNDER THE WALTER RATHBONE BACON TRAVELING SCHOLARSHIP By Edward H. Taylor and Hobart M. Smith INTRODUCTION By tenure of the Walter Rathbone Bacon Traveling Scholarship from 1938 to 1940, the junior author was enabled to continue field studies that had been under way several years on the herpetofauna of Mexico. Aided by his wife, he accumulated a collection of reptiles and amphibians the study of which still continues. A brief summary of the snakes and crocodiles has appeared previously (Smith, 1943). With the aid of the senior author a summary of the amphibians has been completed and forms the basis of the present paper. The liz- ards are being studied as time permits, and a summary of them is contemplated. No survey of the turtles is envisioned. The itin- erary and list of localities visited by the collectors will accompany a later report. The amphibians comprise 10,370 specimens, or about half the total number of herpetological specimens obtained. They represent 27 genera and 146 forms. Thirty-three of the species were undescribed at the time of collecting; the specimens of them secured have formed the basis at least in part for their subsequent descriptions. Eleven of the 33 are represented only by paratypes, while 22 are represented by holotypes. Of the latter, eight are described in the present paper, while all others were described by Taylor (1940c, 1941b, d, e, 1942a-d, 1943a, b) or Smith (1939). -

Evolutionarystudies of Salamanders
AN INTEGRATED APPROACH TO EVOLUTIONARYSTUDIES OF SALAMANDERS DAVIDB. WAKE* CONTENTS der Caudata. Studying organisms from one Introduction ...................................................... 163 perspective can inform and direct investiga- Phylogenetics .................................................... 163 tions that have other goals, and an integra- Hierarchical Approaches .................................. 166 tive approach to organismal evolution can Ontogeny and Phylogeny ................................. 168 result. As evolutionary biologists, our ultimate Recognition of Species ...................................... 170 goal is to understand how biological diversifi- Species Diversity .............................................. 172 cation occurs, and as herpetologists our pri- A Plea for Conservation .................................... 174 mary focus is the whole organism, as exempli- Acknowledgments ............................................ 175 fied by amphibians and reptiles. The examples Literature Cited ............................................... 175 I use are mainly from my own laboratory, be- cause I can see most clearly in these cases how Abstract: This essay is an examination of the ways integration can be achieved. Although this in which seemingly disparate studies focused on a paper is topical in nature, I attempt to provide single, rather small taxon, the Order Caudata, can connections between topics. provide a foundation for development of programs in evolutionary biology. Topics addressed include Any