The Mononuclear Phagocyte System of the Mouse Defined By
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

TEE UNIVERSITY of OKLAHOMA GRADUATE Coiiiege ' A
TEE UNIVERSITY OF OKLAHOMA GRADUATE COIIiEGE ' A COMPARATIVE HISTOLOGICAL AND HISTOCHEMICAL STUDY OF THE ADRENAL GLANDS OF NATIVE RABBITS A THESIS SUBMITTED TO THE GRADUATE FACULTY ±n partial fiolflllment of the requirements for the degree of DOCTOR OF PHILOSOPHY BY I. ERNEST GONZALEZ Oklahoma City, Oklahoma 1955 A COMPARATIVE HISTOLOGICAL AND HISTOCHEMICAL STUDY OF THE ADRENAL GLANDS OF NATIVE RABBITS APEROVED BY THESIS COMMIT' ACKN0WEE33GEMENT The writer wishes to express his profound appreciation and sincere thanks to Dr. Kenneth M. Richter, Department of Anatomy, University of Oklahoma Medical School, for his valuable time, coopera- I _ ition, helpful criticisms, and timely suggestions during the course of j I this investigation; to Dr. Ernest Lachman, Chairman of the Department of Anatomy, for his encouragement and cooperation; to Dr. Garman Daron, Professor of Anatomy, for his many helpful suggestions % and to the University of Oklahoma for a University scholarship. Many other persons have cooperated indirectly in making this investigation possible, and the writer would like to acknowledge also the assistance of Dr. C. Lynn Hayward and Dr. D. Eldon Beck, Depart ment of Zoology, Brigham Young University, for procuring and identify ing many of the native rabbit species; of Mr. Ernest Reiser for his advice during the preparation of the graphic models; and of Mr. Neil Woodward for his assistance with the photomicrographic reproductions. ill TABLE OF CONTENTS Page CHAPTER I 1 Introduction CHAPTER II Materials and MetHods CHAPTER III Observations .............................. 7 Ocbbtona princeps ............»...... ...... 7 Pexicapsular tissue^ capsule, and stroma 7 Vasculature . ......... ............. 8 Innervation............... .... ........ 9 Cortex ........ , . ... ........ ... .. 9 Zona glomerulosa....... ............ 9 Zona fasciculata ............. -

The Morphology, Androgenic Function, Hyperplasia, and Tumors of the Human Ovarian Hilus Cells * William H
THE MORPHOLOGY, ANDROGENIC FUNCTION, HYPERPLASIA, AND TUMORS OF THE HUMAN OVARIAN HILUS CELLS * WILLIAM H. STERNBERG, M.D. (From the Department of Pathology, School of Medicine, Tulane University of Louisiana and the Charity Hospital of Louisiana, New Orleans, La.) The hilus of the human ovary contains nests of cells morphologically identical with testicular Leydig cells, and which, in all probability, pro- duce androgens. Multiple sections through the ovarian hilus and meso- varium will reveal these small nests microscopically in at least 8o per cent of adult ovaries; probably in all adult ovaries if sufficient sections are made. Although they had been noted previously by a number of authors (Aichel,l Bucura,2 and von Winiwarter 3"4) who failed to recog- nize their significance, Berger,5-9 in 1922 and in subsequent years, pre- sented the first sound morphologic studies of the ovarian hilus cells. Nevertheless, there is comparatively little reference to these cells in the American medical literature, and they are not mentioned in stand- ard textbooks of histology, gynecologic pathology, nor in monographs on ovarian tumors (with the exception of Selye's recent "Atlas of Ovarian Tumors"10). The hilus cells are found in clusters along the length of the ovarian hilus and in the adjacent mesovarium. They are, almost without excep- tion, found in contiguity with the nonmyelinated nerves of the hilus, often in intimate relationship to the abundant vascular and lymphatic spaces in this area. Cytologically, a point for point correspondence with the testicular Leydig cells can be established in terms of nuclear and cyto- plasmic detail, lipids, lipochrome pigment, and crystalloids of Reinke. -

The Adrenal Capsule Is a Signaling Center Controlling Cell Renewal and Zonation Through Rspo3
Downloaded from genesdev.cshlp.org on September 24, 2021 - Published by Cold Spring Harbor Laboratory Press RESEARCH COMMUNICATION The permanent cortex is formed through recruitment of The adrenal capsule is a capsular cells in a process that involves SHH signaling signaling center controlling (King et al. 2009). By E17.5, steroidogenic cells have adopted specific expression profiles, with the outermost cell renewal and zonation cell layers (zona glomerulosa [ZG]) producing enzymes Rspo3 that are required for mineralocorticoid production (e.g., through CYP11B2), and deeper layers (zona fasciculata [ZF]) Valerie Vidal,1,2,3,9 Sonia Sacco,1,2,3,9 expressing genes involved in glucocorticoid synthesis Ana Sofia Rocha,1,2,3,8 Fabio da Silva,1,2,3 (Cyp11b1). In humans, but not rodents, a third layer (zona reticularis) can be distinguished that produces an- Clara Panzolini,1,2,3 Typhanie Dumontet,4,5 1,2,3 6 drogens and is located close to the medulla. Several lines Thi Mai Phuong Doan, Jingdong Shan, of evidence suggest that β-catenin plays an important Aleksandra Rak-Raszewska,6 Tom Bird,7 role in adrenal zonation and maintenance. Activation of Seppo Vainio,6 Antoine Martinez,4,5 the β-catenin pathway is restricted to the ZG (Kim et al. and Andreas Schedl1,2,3 2008; Walczak et al. 2014), and ectopic expression leads to the activation of ZG markers in ZF cells (Berthon 1Institute of Biology Valrose, Université de Nice-Sophia, F-06108 et al. 2010). Moreover, β-catenin seems to bind to and con- Nice, France; 2UMR1091, Institut National de la Santé et de la trol the expression of At1r, a gene specifically expressed Recherche Médicale, F-06108 Nice, France; 3CNRS, UMR7277, within the ZG (Berthon et al. -

Histogenesis of Suprarenal Glands at Different Gestational Age Groups
ORIGINAL ARTICLE ASIAN JOURNAL OF MEDICAL SCIENCES Histogenesis of suprarenal glands at different gestational age groups Ravindra Kumar Boddeti1, Subhadra Devi Velichety2 1Lecturer, 2Professor and Head, Department of Anatomy, Sri Padmavathi Medical College for Women, Sri Venkateswara Institute of Medical Sciences, SVIMS University, Tirupathi, Andhra Pradesh, India Submitted: 22-02-2019 Revised: 10-03-2019 Published: 01-05-2019 ABSTRACT Background: The human foetal suprarenal gland is structurally variant from its adult Access this article online counterpart. The most distinctive features of human foetal suprarenal gland and histologically Website: unique foetal zone, was described first by Elliott and Armour in 1911. After the first trimester, the centrally located foetal zone accounts for most of the foetal adrenal mass. The outer zone http://nepjol.info/index.php/AJMS of the foetal suprarenal gland is called the “definitive zone or neo cortex”; this zone likely DOI: 10.3126/ajms.v10i3.22820 gives rise to the adult adrenal glomerulosa. A third zone called “transitional zone”, lies just E-ISSN: 2091-0576 2467-9100 between the neocortex and foetal zone and is believed to develop into the zona fasciculata. P-ISSN: Aims and Objectives: The current study was designed to study the histogenesis of suprarenal glands at different gestational age groups. Materials and Methods: Twenty-eight formalin preserved dead embryos and foetuses of both sexes, were obtained from the Govt. Maternity Hospital & S.V.Medical College, Tirupati, Andhra Pradesh, India. Specimens were grouped according to their gestational age groups (A,B,C,D) A= 0-12 weeks, B= 13-24 weeks, C= 25-36 weeks and D= more than 36 weeks of gestation. -

Pig Gonads, Adrenal Glands and Brain C
Immunoreactive cytochrome P-45017\g=a\in rat and guinea- pig gonads, adrenal glands and brain C. Le Goascogne1, N. Sanan\l=e'\s1, M. Gou\l=e'\zou1, S. Takemori2, S. Kominami2, E. E. Baulieu1 and P. Robel1 1INSERM U33, Communications Hormonales, and Faculté de Médecine, Université Paris-sud, Lab Hormones F-94275 Bicêtre Cedex France 2 Faculty of Integrated Arts and Sciences, Hiroshima University, Hiroshima 730, Japan Summary. The cytochrome P-45017\g=a\-hydroxylase, 17\ar=r\20lyase (P-45017\g=a\) is the key enzyme responsible for the biosynthesis of androgens in steroidogenic organs. Its cellular localization has been examined with an immunohistochemical technique. In immature rat ovary, P-45017\g=a\was first detected in sparse interstitial cells on postnatal Day 8. The number of immunoreactive interstitial cells increased thereafter and the intensity of P-45017\g=a\staining in these cells was highest at 3 weeks of age. The intensity of staining then started to decline and was very faint at Day 35. From 6 weeks on, the distribution of immunoreactive P-45017\g=a\was of the adult type: it was detected exclusively in the thecal cells of the large antral, preovulatory, follicles. P-45017\g=a\was not detectable during pregnancy except on the day of parturition, when thecal cells were transiently immunoreactive. The staining had vanished 24 h after delivery. Human chorionic gonadotrophin (hCG), injected into immature females on Days 24 to 26, induced P-45017\g=a\prematurely in thecal cells. When injected on Days 12 to 14 of pregnancy, hCG also induced P-45017\g=a\in the thecal cells surrounding the largest follicles, whereas the interstitial and luteal cells were not immunostained. -

Adrenal Gland Hormones
CHAPTER 8 Adrenal Gland Hormones Devra K. Dang, PharmD, BCPS, CDE, FNAP | Trinh Pham, PharmD, BCOP | Jennifer J. Lee, PharmD, BCPS, CDE LEARNING OBJECTIVES KEY TERMS AND DEFINITIONS After completing this chapter, you should be able to ACTH (adrenocorticotropic hormone) — a hormone produced 1. Identify the hormones produced by the adrenal glands by the pituitary gland that stimulates 2. Describe the functions of mineralocorticoids and glucocorticoids in the body the adrenal cortex to produce glucocorticoids, mineralocorticoids, 3. Recognize the signs and symptoms of adrenal insuffi ciency and androgens. PART 4. Describe the pharmacological treatment of patients with acute and chronic adrenal Addison ’ s disease — a disorder insuffi ciency in which the adrenal glands do not produce enough steroid hormones. 3 5. Recognize the signs and symptoms of Cushing ’ s syndrome and the result of too Adenoma — a benign much cortisol (noncancerous) tumor of glandular 6. Describe the pharmacologic and nonpharmacologic management of patients with origin. Cushing ’ s syndrome Adrenal insuffi ciency — a term 7. List management strategies for administration of glucocorticoid and mineralocorti- referring to a defi ciency in the levels of adrenal hormones. coid therapy to avoid development of adrenal disorders Aldosterone — the hormone produced by the adrenal glands that regulates the balance of sodium, he adrenal glands are an integral part of the endocrine system, secreting water, and potassium concentrations in the body. T hormones that act throughout the body to regulate functions and promote Corticotropin-releasing homeostasis. In addition to the neurotransmitters epinephrine and norepineph- hormone (CRH) — a hormone rine, the corticosteroids secreted by the adrenal glands are vital to a wide released by the hypothalamus that variety of physiological processes. -

Hypothalamushypothalamus -- Pituitarypituitary -- Adrenaladrenal Glandsglands
HypothalamusHypothalamus -- pituitarypituitary -- adrenaladrenal glandsglands Magdalena Gibas-Dorna MD, PhD Dept. of Physiology University of Medical Sciences Poznań, Poland Hypothalamus - general director of the hormone system. At every moment, the hypothalamus analyses messages coming from: the brain and different regions of the body. Homeostatic functions of hypothalamus include maintaining a stable body temperature, controlling food intake, controlling blood pressure, ensuring a fluid balance, and even proper sleep patterns. Cell bodies of neurons that produce releasing/inhibiting hormones Hypothalamus HypothalamusHypothalamus releases Arterial flow Primary capillaries in median eminence hormones at Long Releasing Portal hormones Anterior veins median eminence pituitary hormone Releasing/ inhibiting hormones and sends to anterior pituitary ANTERIOR PITUITARY via portalportal veinvein. Secretory cells that produce anterior pituitary hormones Anterior pituitary hormones Venous outflow Gonadotropic Thyroid- Proactin hormones stimulating ACTH Growth (FSH and LH) hormone hormone ControlControl ofof pituitarypituitary hormonehormone secretionsecretion byby hypothalamushypothalamus • Secretion by the anterioranterior pituitarypituitary is controlled by hormones called hypothalamic releasing hormones and inhibitory hormones conducted to the anterior pituitary through hypothalamichypothalamic -- hypophysialhypophysial portalportal vesselsvessels .. • PosteriorPosterior pituitarypituitary secrets two hormones, which are synthesized within cell -

Adrenal Gland
ADRENAL GLAND Objectives: ◧ Editing file • Differentiate between adrenal cortex ◧ Important and medulla. ◧ Doctor notes / Extra • Identify the histological features of each cortical zone and its cells. • Identify the histological features of the medullary cells. 438 Histology Team Endocrine Block Zona glomerulosa Stroma Cortex Zona fasciculata Parenchyma Zona reticularis Adrenal gland Adrenal Medulla The adrenal cortex layers have 5 features in common: extra but important 1. Suprarenal artery 1- Acidophilic cytoplasm 2. Capsule 2- Abundant SER 3. Zona glomerulosa 3- Numerous mitochondria 4. Zona fasciculata 5. Zona reticularis 4- Mitochondrial cristae is tubular 6. Medulla 5- few Droplet of lipids ( expect Zona fasciculata rich in lipids) 7. Central vein of medulla 438 Histology Team - Endocrine Block 2 Adrenal Cortex Zona fasciculata Zona glomerulosa Zona reticularis (spongiocytes) ● formed of clusters of small • It is the intermediate and the largest •It is the innermost layer of adrenal columnar cells that are rich in layer of the cortex. cortex. SER and mitochondria. • It is formed of columns of large •It is formed of anastomosing ● Produces mineralocorticoids polyhedral cells that are separated cords of deep acidophilic cells. e.g. aldosterone hormone by longitudinal sinusoidal capillaries. •Its cells contains few lipofuscin (Reabsorb all the remaining • Its cells are rich in lipids, so they and lipid droplets. sodium, and passively the appear empty in sections chloride, from the lumen of the (spongiocytes). •The cells secrete androgens. distal renal tubules into the renal • Its cells are rich in mitochondria (with interstitium. In addition, tubular cristae),SER and lipofuscin potassium and hydrogen ions pigments. are actively secreted into the lumen). -

Steroidogenic Differentiation and PKA Signaling Are Programmed by Histone Methyltransferase EZH2 in the Adrenal Cortex
Steroidogenic differentiation and PKA signaling are programmed by histone methyltransferase EZH2 in the adrenal cortex Mickael Mathieua, Coralie Drelona,1, Stéphanie Rodrigueza,1, Houda Tabbala, Amandine Septiera, Christelle Damon-Soubeyranda, Typhanie Dumonteta, Annabel Berthona,b, Isabelle Sahut-Barnolaa, Cyril Djaria, Marie Batisse-Ligniera,c, Jean-Christophe Pointuda, Damien Richardd, Gwenneg Kerdivele, Marie-Ange Calméjanee, Valentina Boevae, Igor Tauverona,c, Anne-Marie Lefrançois-Martineza, Antoine Martineza, and Pierre Vala,2 aGénétique Reproduction & Développement, CNRS UMR 6293, Inserm U1103, Université Clermont Auvergne, 63001 Clermont-Ferrand, France; bEunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20892-1103; cService d’Endocrinologie, Faculté de Médecine, Centre Hospitalier Universitaire, Université Clermont Auvergne, 63000 Clermont-Ferrand, France; dService de Pharmacologie Médicale, UMR Inserm 1107 Neuro-Dol, Centre Hospitalier Universitaire, Université Clermont Auvergne, 63000 Clermont-Ferrand, France; and eInstitut Cochin, Inserm U1016, CNRS, UMR 8104, Université Paris Descartes UMR-S1016, 75014 Paris, France Edited by Michael G. Rosenfeld, University of California, San Diego, La Jolla, CA, and approved November 15, 2018 (received for review May 29, 2018) Adrenal cortex steroids are essential for body homeostasis, and tribute to the pool of capsular GLI1-positive, SF1-negative pro- adrenal insufficiency is a life-threatening condition. Adrenal endo- genitors that were found during embryonic development but also in crine activity is maintained through recruitment of subcapsular the adult (9). Altogether, these data raise the question of the mo- progenitor cells that follow a unidirectional differentiation path from lecular mechanisms that ensure that progenitors acquire and zona glomerulosa to zona fasciculata (zF). -
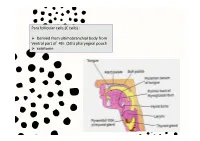
Para Follicular Cells (C Cells)
Para follicular cells (C cells) : Derived from ultimobranchial body from Ventral part of 4th (5th) pharyngeal pouch calcitonin Thyroglossal cyct: path of thyroid descending Position of occur: Inf. To the body of hyoid 50% Base of tongue Close to thyroid cartilage Thyroglossal fistula Abberant thyroid tissue: path of thyroid descending Base of tongue Histology of the thyroid gland Thyroid gland: parafollicular cells: Capsule / trabeculae Follicles / reticular fiber / Decreased Ca+ in blood by 2 ways: basal lamina Follicular cells/ basal lamina 1. Transport Ca from blood to musculoskeletal system parafollicular cells 2. Prevent bone absorption by osteoclast cells Colloid (hormone storage) Produce energy & temperature for body activity Histology of the thyroid gland Thyroid hormones: Tri-iodothyronin Tetra-iodothyronin Calcitonin Fenestrated capillary Synthesis and secretion of thyroid hormones T3 and T4 Synthesis and secretion of thyroid hormones T3 and T4 Synthesis and mechanism of action of calcitonin Gravies disease (exophthalmic goiter / toxic goiter): excessive amounts of thyroid hormones are released into the circulation detectable levels of autoantibodies abnormal immunoglobulins (IgG) bind to the TSH receptor in-creased thyroid hormone secretion Because of negative feedback, the levels of TSH in the circulation are usually normal Hypertrophy thyroid hormone Is abnormally high range increased metabolism Features: weight loss / excessive sweating / tachycardia /nervousness / protrusion of the eyeballs / retraction of the -

A Histocytological Study of Age Changes in the Canine Adrenal Gland Studied by Light and Electron Microscopy Ronald Loral Hullinger Iowa State University
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1968 A histocytological study of age changes in the canine adrenal gland studied by light and electron microscopy Ronald Loral Hullinger Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Animal Structures Commons, and the Veterinary Anatomy Commons Recommended Citation Hullinger, Ronald Loral, "A histocytological study of age changes in the canine adrenal gland studied by light and electron microscopy" (1968). Retrospective Theses and Dissertations. 3674. https://lib.dr.iastate.edu/rtd/3674 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. This dlsseriaUon has been microfihned exactly as received 68-14,799 HULLINGER, D.V.M., Ronald Loral, 1941- A HISTOCYTOLOGICAL STUDY OF AGE CHANGES IN THE CANINE ADRENAL GLAND STUDIED BY LIGHT AND ELECTRON MICROSCOPY. Iowa State University, Ph,D., 1968 Anatomy University Microfilms, Inc., Ann Arbor, Michigan A HISTOCYTOLOGICAL STUDY OF AGE CHANGES IN THE CANINE ADRENAL GLAND STUDIED BY LIGHT AND ELECTRON MICROSCOPY by ' Ronald Loral Hullinger , D. v,'- A Dissertation Submitted to the Graduate Faculty in Partial Fulfillment of The Requirements for the Degree of DOCTOR OF PHILOSOPHY Major Subject: Veterinary Anatomy Approved; Signature was redacted for privacy. In Charge of Majpr^Work Signature was redacted for privacy. .rtment Signature was redacted for privacy. -

Adrenal Cortical Tumors, Pheochromocytomas and Paragangliomas
Modern Pathology (2011) 24, S58–S65 S58 & 2011 USCAP, Inc. All rights reserved 0893-3952/11 $32.00 Adrenal cortical tumors, pheochromocytomas and paragangliomas Ricardo V Lloyd Department of Pathology, University of Wisconsin School of Medicine and Public Health, Madison, WI, USA Distinguishing adrenal cortical adenomas from carcinomas may be a difficult diagnostic problem. The criteria of Weiss are very useful because of their reliance on histologic features. From a practical perspective, the most useful criteria to separate adenomas from carcinomas include tumor size, presence of necrosis and mitotic activity including atypical mitoses. Adrenal cortical neoplasms in pediatric patients are more difficult to diagnose and to separate adenomas from carcinomas. The diagnosis of pediatric adrenal cortical carcinoma requires a higher tumor weight, larger tumor size and more mitoses compared with carcinomas in adults. Pheochromocytomas are chromaffin-derived tumors that develop in the adrenal gland. Paragangliomas are tumors arising from paraganglia that are distributed along the parasympathetic nerves and sympathetic chain. Positive staining for chromogranin and synaptophysin is present in the chief cells, whereas the sustentacular cells are positive for S100 protein. Hereditary conditions associated with pheochromocytomas include multiple endocrine neoplasia 2A and 2B, Von Hippel–Lindau disease and neurofibromatosis I. Hereditary paraganglioma syndromes with mutations of SDHB, SDHC and SDHD are associated with paragangliomas and some pheochromocytomas.