Implications for Communication
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
M.A. Maciver's Dissertation
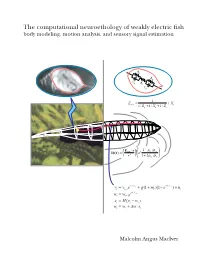
The computational neuroethology of weakly electric fish body modeling, motion analysis, and sensory signal estimation R s R s C R s i C s 1 Z = + X prey + + s 1/ Rp 1/ Xp 1/ Zs −ρ ρ E ⋅r 1 p / w δφ(r) = fish a3 3 + ρ ρ r 1 2 p / w − t − t = 1/ v + + − 1/ v + vt vt−1e g(1 mt )(1 e ) nt − t = 1/ w wt wt−1e = − st H(vt wt ) = + ∆ ⋅ wt wt w st Malcolm Angus MacIver THE COMPUTATIONAL NEUROETHOLOGY OF WEAKLY ELECTRIC FISH: BODY MODELING, MOTION ANALYSIS, AND SENSORY SIGNAL ESTIMATION BY MALCOLM ANGUS MACIVER B.Sc., University of Toronto, 1991 M.A., University of Toronto, 1992 THESIS Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Neuroscience in the Graduate College of the University of Illinois at Urbana-Champaign, 2001 Urbana, Illinois c Copyright by Malcolm Angus MacIver, 2001 ABSTRACT Animals actively influence the content and quality of sensory information they acquire through the positioning of peripheral sensory surfaces. Investigation of how the body and brain work together for sensory acquisition is hindered by 1) the limited number of techniques for tracking sensory surfaces, few of which provide data on the position of the entire body surface, and by 2) our inability to measure the thousands of sensory afferents stimulated during behav- ior. I present research on sensory acquisition in weakly electric fish of the genus Apteronotus, where I overcame the first barrier by developing a markerless tracking system and have de- ployed a computational approach toward overcoming the second barrier. -

Chirping and Asymmetric Jamming Avoidance Responses in the Electric Fish Distocyclus Conirostris Jacquelyn M
© 2018. Published by The Company of Biologists Ltd | Journal of Experimental Biology (2018) 221, jeb178913. doi:10.1242/jeb.178913 SHORT COMMUNICATION Chirping and asymmetric jamming avoidance responses in the electric fish Distocyclus conirostris Jacquelyn M. Petzold1,2, JoséA. Alves-Gomes3 and G. Troy Smith1,2,* ABSTRACT of two of more EODs creates a periodic amplitude modulation Electrosensory systems of weakly electric fish must accommodate (beat). Beat frequency is equal to the difference between the EOD competing demands of sensing the environment (electrolocation) frequencies (EODfs) of the two interacting fish. Fish use the beat and receiving social information (electrocommunication). The and the relative geometry of the interacting signals to estimate jamming avoidance response (JAR) is a behavioral strategy thought conspecific EODfs, which convey important social information to reduce electrosensory interference from conspecific signals close (Smith, 2013; Dunlap, et al., 2017). However, slow beats (<10 Hz) in frequency. We used playback experiments to characterize electric created by interactions between fish with similar EODfs can impair organ discharge frequency (EODf), chirping behavior and the JAR of the electrolocation function of the EOD by masking localized EOD Distocyclus conirostris, a gregarious electric fish species. EODs of D. distortions (Heiligenberg, 1973; Matsubara and Heiligenberg, conirostris had low frequencies (∼80–200 Hz) that shifted in response 1978). The JAR is a stereotyped response in which an electric to playback stimuli. Fish consistently lowered EODf in response to fish increases or decreases its EODf to increase beat frequency and higher-frequency stimuli but inconsistently raised or lowered EODf in thereby reduce or eliminate the interference caused by slow beats response to lower-frequency stimuli. -

Hormones and Sexual Behavior of Teleost Fishes
Chapter 7 Hormones and Sexual Behavior of Teleost Fishes y David M. Gonc¸alves*, and Rui F. Oliveira*,** y * Instituto Superior de Psicologia Aplicada, Lisboa, Portugal, Universidade do Algarve, Faro, Portugal, ** Instituto Gulbenkian de Cieˆncia, Oeiras, Portugal more variable during the initial stages of the sequence and SUMMARY more stereotyped towards its end. To account for this Fishes are an excellent group for studying the mechanisms through which hormones modulate the expression of sexual variation, these researchers suggested that an initial appe- behaviors in vertebrates. First, they have radiated virtually titive phase, defined as the phase of searching towards the throughout all aquatic environments and this is reflected in an goal, can be distinguished from a final consummatory extraordinary diversity of mating systems and reproductive phase, defined as the stage when the goal is reached behaviors. Second, many species present a remarkable plasticity (Sherrington, 1906; Craig, 1917). Although this distinction in their sexual displays, as exemplified by fishes that change sex or is still widely applied in studies investigating the mecha- that adopt more than one reproductive tactic during their lifetime, nisms of behavior, there is an ongoing debate on the and this plasticity seems to be mediated by hormones. Third, the usefulness of these terms. In a recent review, Sachs (2007) fish neuroendocrine system is well conserved among vertebrates identified some problems in the current use of the and the mechanisms of hormonal action in behavior are likely to appetitive/consummatory dichotomy. These include the share similarities with those of other vertebrates. We review the difficulties in defining the boundary between the two pha- role of hormones and neuropeptides in the modulation of fish sexual displays. -

JAGE-691 Fish Cognition and Consciousness Colin Allen [email protected] Phone
JAGE-691 Fish Cognition and Consciousness Colin Allen [email protected] phone: +1-812-606-0881 fax: +1-812-855-3631 Program in Cognitive Science and Department of History and Philosophy of Science Indiana University, Bloomington, IN 47405 USA ABSTRACT. Questions about fish consciousness and cognition are receiving increasing attention. In this paper, I explain why one must be careful to avoid drawing conclusions too hastily about this hugely di- verse set of species. Keywords. Fish, learning, cognition, consciousness 1. Introduction to the controversy The cognitive and mental capacities of fish are a current topic of scientific controversy, and consciousness is the most contentious of topics. In a recent review article, Michel Cabanac and coauthors (Cabanac et al. 2009) argue that consciousness did not emerge until the early Amniota, the group of species that includes mammals, birds, and "reptiles.” The latter term is in scare quotes because biologists consider it a paraphy- letic group (i.e., a group that contains just a subset of the descendants of its common ancestor) that is im- proper for classification purposes due to its exclusion of the birds, which descended from the saurians. Amniotes are characterized by an embryonic membrane that makes terrestrial reproduction feasible. The amphibians, lacking this adaptation, are constrained to place their eggs in an aqueous environment for proper development. These biological details are important because of the nature of some of the evidence that Cabanac et al. bring to bear on the question of consciousness in fish – evidence that I shall maintain seems skewed towards other adaptations that have to do with terrestrial life. -

Effects of Restraint and Immobilization on Electrosensory Behaviors of Weakly Electric Fish
Effects of Restraint and Immobilization on Electrosensory Behaviors of Weakly Electric Fish Éva M. Hitschfeld, Sarah A. Stamper, Katrin Vonderschen, Eric S. Fortune, and Maurice J. Chacron Abstract Introduction Weakly electric fi shes have been an important model system eakly electric fi sh emit an electric organ dis- in behavioral neuroscience for more than 40 years. These charge (EOD1) using a specialized electric or- fi shes use a specialized electric organ to produce an electric W gan located in the tail. EOD properties such as fi eld that is typically below 1 volt/cm and serves in many be- fundamental frequency are highly regulated by central ner- haviors including social communication and prey detection. vous system (CNS) circuits (Heiligenberg 1991). Electrore- Electrical behaviors are easy to study because inexpensive ceptors in the animal’s skin detect perturbations of the EOD and widely available tools enable continuous monitoring of caused by nearby objects, prey, and the electric fi elds of con- the electric fi eld of individual or groups of interacting fi sh. specifi cs (Nelson and MacIver 1999; Turner et al. 1999; Weakly electric fi sh have been routinely used in tightly con- Zakon et al. 2002). For many species of electric fi sh, simple trolled neurophysiological experiments in which the animal is sine wave electric signals at species-appropriate frequencies immobilized using neuromuscular blockers (e.g., curare). Al- and amplitudes can elicit behavioral responses from conspe- though experiments that involve immobilization are generally cifi cs (Heiligenberg 1991). discouraged because it eliminates movement-based behav- Most species of weakly electric fi shes actively modulate ioral signs of pain and distress, many observable electrosen- their EODs depending on the behavioral context. -

Electrocommunication for Weakly Electric Fish
Electrocommunication for weakly electric fish A. Scapin Research Report No. 2019-14 March 2019 Seminar für Angewandte Mathematik Eidgenössische Technische Hochschule CH-8092 Zürich Switzerland ____________________________________________________________________________________________________ Funding SNF: 200021-172483 ELECTROCOMMUNICATION FOR WEAKLY ELECTRIC FISH ANDREA SCAPIN Abstract. This paper addresses the problem of the electro-communication for weakly electric fish. In particular we aim at sheding light on how the fish circumvent the jamming issue for both electro-communication and active electro-sensing. A real-time tracking algorithm is presented. 1. Introduction In this paper we address the problem of studying the behaviour of two weakly electric fish when they populate the same environment. Those kind of animals orient themselves at night in complete darkness by using their active electro-sensing system. They gen- erate a stable, relatively high-frequency, weak electric field and transdermically perceive the corresponding feedback by means of many receptors on its skin. Since they have an electric sense that allows underwater navigation, target classification and intraspecific communication, they are privileged animals for bio-inspiring man-built autonomous sys- tems [14, 18, 19, 21, 24, 29, 39, 40]. For electro-communication purposes, in processing sensory information, this system has to separate feedback associated with their own signals from interfering sensations caused by signals from other animals. Wave and pulse species employ different mechanisms to minimize interference with EODs of conspecifics. It has been observed that certain wave species, having wave-type electric organ discharge (EOD) waveforms, such as Eigenmannia and Gymnarchus, reflexively shift their EOD frequency away from interfering frequencies of nearby conspecifics, in order to avoid “jamming” each others electrical signals. -

The Evolution of Echolocation for Predation
Symp. zool. Soc. Land. (1993) No. 65: 39-63 The evolution of echolocation for predation J. R. SPEAKMAN DepartmentofZoowgy University of Aberdeen Aberdeen AB9 2TN, UK Synopsis Active detection of food items by echolocation has some obvious advantages over passive detection, since it affords independence from ambient light and sound levels. For predatory animals, however, echolocation would also appear to have a significant disadvanrage-i-the echolocation calls might alert prey to the predator's presence. Surprisingly, therefore, all but two of the different groups of vertebrates that have evolved echolocation are predatory. Despite the diversity of predatory taxa in which echolocation has evolved it is still a relatively uncommon form of perception. It has been suggested that a major constraint on the evolution of echolocation is its high energy cost, due to rapid attenuation of sound in air. The cost of producing echolocation calls has been measured in insectivorous bats, whilst hanging at rest. These measures confirm that echolocation is extremely costly. However, bats normally echolocate in flight which also has a high cost. How bats cope with the high cost of echolocation, when it is combined with flight, is therefore of extreme interest. Measures of the energy cost of flight of small echolocating bats ggest that the cost is no greater than that for non-echolocating birds and bats. The son for this apparent economy is that the same muscles which flap the wings also tilate the lungs, and produce the pulse of breath which generates the echolocation For a bat in flight, therefore, the additional cost of echolocating is very low, sr for a bat on the ground, and presumably other terrestrial vertebrates, the cost ry high. -

A New and Improved Electric Fish Finder with Resources for Printed Circuit Board Fabrication
Neotropical Ichthyology, 17(4): e190099, 2019 Journal homepage: www.scielo.br/ni DOI: 10.1590/1982-0224-20190099 Published online: 25 November 2019 (ISSN 1982-0224) Copyright © 2019 Sociedade Brasileira de Ictiologia Printed: 13 December 2019 (ISSN 1679-6225) Original article A new and improved electric fish finder with resources for printed circuit board fabrication Michael A. Haag, Jeffrey C. Lambert, Joseph C. Waddell and William G.R. Crampton We describe the circuit design, construction, and operation of a field-portable electric fish finder (an AC-coupled wide- band differential bio-amplifier with loudspeaker output). This device permits detection and monitoring of the electric organ discharges generated by neotropical gymnotiform fishes (as well as the mormyroid fishes of tropical Africa). Our design is modified from earlier versions to optimize detection performance and stability over a wider range of ambient water conductivity, including under conditions of extremely low conductivity (< ca. 10 μScm-1). Our new electric fish finder design also incorporates complete waterproofing and longer battery autonomy. We provide Gerber and Eagle files made with the electronic design automation software ‘Autodesk Eagle’ to allow researchers to order printed circuit boards directly from commercial manufacturers. Keywords: Bio-amplifier, Electric organ discharge, Electroreception, Gymnotiformes, Mormyridae. Nós descrevemos o projeto de circuitos eletrônicos e as instrucões para a construção e uso de um detector de peixes elétricos portátil (bio-amplificador diferencial de banda-larga com acoplamento AC). Este aparelho permite a detecção e o monitoramento das descargas de órgãos elétricos gerados por peixes neotropicais da ordem Gymnotiformes (assim como dos peixes mormirídeos da África Tropical). -

Mormyridae, Teleostei)
Ethology 103, 404-420 (1997) © 1997 Blackwell Wissenschafts-Verlag, Berlin ISSN 0179-1613 Zoologisches Institut der Universitdt Regensburg, Regensburg Electrocommunication and Social Behaviour in Marcusenius senegalensis (Mormyridae, Teleostei) ANDREAS SCHEFFEL & BERND KRAMER SCHEFFEL, A. & KRAMER, B. 1997: Electrocommuncation and social behaviour in Marcusenius senegalensis (Mormyridae, Teleostei). Ethology 103, 404—420. Abstract The electric organ discharges (EODs) of Marcusenius senegalensis, a West African freshwater fish, are bipolar pulses of short duration (220 ± SE 13 µs). In males (n = 10; 10.1-13.1 cm standard length-which is around the si2e of getting mature), the duration of EOD pulses was of significantly greater variance than in females (n = 9; 9.8—12.8 cm standard length). Male EODs also showed a tendency for a longer duration than female EODs. Groups of three as well as of 14 M. senegalensis formed temporary schools in a 'naturally' equipped 720-1 tank. While swimming slowly in a loose school during their nocturnal active phase, fish discharged in irregular long—short—long inter-EOD interval patterns. Near neighbours displayed a tendency to discharge in intervals of similar duration (nearest neighbour distance < 1/2 fish length). On removal of a plastic partition that had separated a pair of fish for at least 3 days, mutual threat displays followed by fighting were observed. During threatening, the fish alternated regularly between bursts of a high discharge rate and short discharge breaks; the rate of change was 4/s. The subdominant animal in a group of two was attacked frequently and often ceased discharging when the dominant fish approached. -

Evidence for Mutual Allocation of Social Attention Through Interactive Signaling in a Mormyrid Weakly Electric Fish
Evidence for mutual allocation of social attention through interactive signaling in a mormyrid weakly electric fish Martin Worma,1, Tim Landgrafb, Julia Prumea, Hai Nguyenb, Frank Kirschbaumc, and Gerhard von der Emdea aInstitut für Zoologie, Neuroethologie/Sensorische Ökologie, Universität Bonn, 53115 Bonn, Germany; bInstitut für Informatik, Fachbereich Informatik und Mathematik, Freie Universität Berlin, 14195 Berlin, Germany; and cBiologie und Ökologie der Fische, Lebenswissenschaftliche Fakultät, Humboldt-Universität-zu Berlin, 10115 Berlin, Germany Edited by John G. Hildebrand, University of Arizona, Tucson, AZ, and approved May 16, 2018 (received for review January 26, 2018) Mormyrid weakly electric fish produce electric organ discharges responses from a conspecific. We solved both problems by using (EODs) for active electrolocation and electrocommunication. These a freely moving robotic fish capable of emitting either predefined pulses are emitted with variable interdischarge intervals (IDIs) or dynamic sequences of playback EODs in an interactive be- resulting in temporal discharge patterns and interactive signaling havioral experiment with single individuals of the weakly electric episodes with nearby conspecifics. However, unequivocal assign- fish Mormyrus rume proboscirostris. ment of interactive signaling to a specific behavioral context has Robotic fish have been successfully employed to investigate the proven to be challenging. Using an ethorobotical approach, we features determining attraction between individual fish (14–16), as confronted single individuals of weakly electric Mormyrus rume well as collective decision making and internal dynamics in groups – proboscirostris with a mobile fish robot capable of interacting of fish in shoals (17 22). Similar experiments have demonstrated both physically, on arbitrary trajectories, as well as electrically, that mormyrids are attracted to a mobile fish replica playing back by generating echo responses through playback of species- electric signals (23, 24). -

In Praise of Fishes: Précis of What a Fish Knows (Balcombe 2016)
Balcombe, Jonathan (2016) In praise of fishes: Précis of What a fish knows (Balcombe 2016). Animal Sentience 8(1) DOI: 10.51291/2377-7478.1112 This article has appeared in the journal Animal Sentience, a peer-reviewed journal on animal cognition and feeling. It has been made open access, free for all, by WellBeing International and deposited in the WBI Studies Repository. For more information, please contact [email protected]. Animal Sentience 2016.095: Balcombe on Fishes Know [Editorial Note: Multiple invited reviews of this book will be appearing in upcoming issues of Animal Sentience, together with the author’s Response.] Multiple Book Review: Animal Sentience publishes multiple reviews of selected books whose Précis is published in the journal along with the reviews and the author’s response(s). Most of the multiple book reviews are invited individually by the Editor but freely submitted reviews of the selected books whose Précis has appeared can also be considered for publication: Please write to the editor to request a review copy. Books can also be nominated and reviewers can be proposed by writing to the Editor. Instructions: http://animalstudiesrepository.org/animsent/guidelines.html In praise of fishes Précis of What a fish knows (Balcombe 2016) Jonathan Balcombe Humane Society Institute for Science and Policy The Humane Society of the United States Abstract: Our relationship to fishes in the modern era is deeply problematic. We kill and consume more of them than any other group of vertebrates. At the same time, advances in our knowledge of fishes and their capabilities are gaining speed. -

12 Electroreception and Electrogenesis
2022_C012.fm Page 431 Tuesday, June 7, 2005 4:03 PM Electroreception and 12 Electrogenesis James S. Albert and William G.R. Crampton CONTENTS I. Introduction to Electroreception and Electrogenesis ........................................................432 II. Phylogeny of Electroreception and Electrogenesis...........................................................433 III. Passive Electroreception ....................................................................................................437 A. Electroreception and Mechanoreception: Vertebrate Laterosensory Systems..........438 B. Electroreception in Lampreys....................................................................................438 C. Electroreception in Gnathostomes.............................................................................439 1. Electroreception in Chondrichthyans ..................................................................439 2. Passive Electroreception in Electric Skates and Rays ........................................440 3. Nonteleost Bony Fishes.......................................................................................441 D. Electroreception in Teleosts ......................................................................................441 1. Passive Electroreception in Weakly Electrogenic Siluriformes..........................442 2. Transition from Electric Communication to Active Electroreception................442 IV. Active Electroreception......................................................................................................443