Lecture. 6 Population Growth – Intraspecific Competition Individuals
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Effective Population Size and Genetic Conservation Criteria for Bull Trout
North American Journal of Fisheries Management 21:756±764, 2001 q Copyright by the American Fisheries Society 2001 Effective Population Size and Genetic Conservation Criteria for Bull Trout B. E. RIEMAN* U.S. Department of Agriculture Forest Service, Rocky Mountain Research Station, 316 East Myrtle, Boise, Idaho 83702, USA F. W. A LLENDORF Division of Biological Sciences, University of Montana, Missoula, Montana 59812, USA Abstract.ÐEffective population size (Ne) is an important concept in the management of threatened species like bull trout Salvelinus con¯uentus. General guidelines suggest that effective population sizes of 50 or 500 are essential to minimize inbreeding effects or maintain adaptive genetic variation, respectively. Although Ne strongly depends on census population size, it also depends on demographic and life history characteristics that complicate any estimates. This is an especially dif®cult problem for species like bull trout, which have overlapping generations; biologists may monitor annual population number but lack more detailed information on demographic population structure or life history. We used a generalized, age-structured simulation model to relate Ne to adult numbers under a range of life histories and other conditions characteristic of bull trout populations. Effective population size varied strongly with the effects of the demographic and environmental variation included in our simulations. Our most realistic estimates of Ne were between about 0.5 and 1.0 times the mean number of adults spawning annually. We conclude that cautious long-term management goals for bull trout populations should include an average of at least 1,000 adults spawning each year. Where local populations are too small, managers should seek to conserve a collection of interconnected populations that is at least large enough in total to meet this minimum. -

Routledge Handbook of Ecological and Environmental Restoration the Principles of Restoration Ecology at Population Scales
This article was downloaded by: 10.3.98.104 On: 26 Sep 2021 Access details: subscription number Publisher: Routledge Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: 5 Howick Place, London SW1P 1WG, UK Routledge Handbook of Ecological and Environmental Restoration Stuart K. Allison, Stephen D. Murphy The Principles of Restoration Ecology at Population Scales Publication details https://www.routledgehandbooks.com/doi/10.4324/9781315685977.ch3 Stephen D. Murphy, Michael J. McTavish, Heather A. Cray Published online on: 23 May 2017 How to cite :- Stephen D. Murphy, Michael J. McTavish, Heather A. Cray. 23 May 2017, The Principles of Restoration Ecology at Population Scales from: Routledge Handbook of Ecological and Environmental Restoration Routledge Accessed on: 26 Sep 2021 https://www.routledgehandbooks.com/doi/10.4324/9781315685977.ch3 PLEASE SCROLL DOWN FOR DOCUMENT Full terms and conditions of use: https://www.routledgehandbooks.com/legal-notices/terms This Document PDF may be used for research, teaching and private study purposes. Any substantial or systematic reproductions, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The publisher shall not be liable for an loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. 3 THE PRINCIPLES OF RESTORATION ECOLOGY AT POPULATION SCALES Stephen D. -

Globalization and Infectious Diseases: a Review of the Linkages
TDR/STR/SEB/ST/04.2 SPECIAL TOPICS NO.3 Globalization and infectious diseases: A review of the linkages Social, Economic and Behavioural (SEB) Research UNICEF/UNDP/World Bank/WHO Special Programme for Research & Training in Tropical Diseases (TDR) The "Special Topics in Social, Economic and Behavioural (SEB) Research" series are peer-reviewed publications commissioned by the TDR Steering Committee for Social, Economic and Behavioural Research. For further information please contact: Dr Johannes Sommerfeld Manager Steering Committee for Social, Economic and Behavioural Research (SEB) UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR) World Health Organization 20, Avenue Appia CH-1211 Geneva 27 Switzerland E-mail: [email protected] TDR/STR/SEB/ST/04.2 Globalization and infectious diseases: A review of the linkages Lance Saker,1 MSc MRCP Kelley Lee,1 MPA, MA, D.Phil. Barbara Cannito,1 MSc Anna Gilmore,2 MBBS, DTM&H, MSc, MFPHM Diarmid Campbell-Lendrum,1 D.Phil. 1 Centre on Global Change and Health London School of Hygiene & Tropical Medicine Keppel Street, London WC1E 7HT, UK 2 European Centre on Health of Societies in Transition (ECOHOST) London School of Hygiene & Tropical Medicine Keppel Street, London WC1E 7HT, UK TDR/STR/SEB/ST/04.2 Copyright © World Health Organization on behalf of the Special Programme for Research and Training in Tropical Diseases 2004 All rights reserved. The use of content from this health information product for all non-commercial education, training and information purposes is encouraged, including translation, quotation and reproduction, in any medium, but the content must not be changed and full acknowledgement of the source must be clearly stated. -

Litter Pollution in Densely Versus Sparsely Populated Areas: Dog River Watershed
LITTER POLLUTION IN DENSELY VERSUS SPARSELY POPULATED AREAS: DOG RIVER WATERSHED Gabrielle M. Hudson, Department of Earth Sciences, University of South Alabama, Mobile, AL 36688. E-mail: [email protected]. It is commonly known that when humans populate an area that area is usually subject to some environmental degradation. One of the more common aspects of environmental degradation is litter. This type of degradation is no stranger to the Dog River watershed. For years residents have seen the rivers in this watershed covered in trash, specifically after rain events. The vast majority of the trash is a result of litter from roadsides being carried into the river via drainage pipes. This paper is a comparative study of litter in areas of varying population densities in the Dog River watershed. It seeks to distinguish between the amount of litter found in densely populated areas and sparsely populated areas, and to find out if there is a correlation between population density and litter. I utilize GIS to map population density of the Dog River watershed, and analyze and compare the amounts of litter in areas of sparse and dense populations. The results show that there is no correlation between population density and litter. It also shows that there is no difference in the amounts of litter found in densely and sparsely populated areas. Keyword: litter, population density, watershed Introduction Pollution has long been an issue in the Dog River watershed, in particular litter pollution. The extent of the pollution has not gone unnoticed. There are groups of people and organizations who have taken increased interest in the Dog River watershed with intentions of reducing pollution, including Dog River Clearwater Revival and its numerous volunteers. -

Population Growth & Resource Capacity
Population Growth & Resource Capacity Part 1 Population Projections Between 1950 and 2005, population growth in the U.S. has been nearly linear, as shown in figure 1. Figure 1 U.S. Population in Billions 0.4 0.3 0.2 Actual Growth 0.1 Linear Approximation - - - H L Year 1950 1960 1970 1980 1990 2000 2010 Source: Population Division of the Department of Economic and Social Affairs of the United Nations Secretariat. World Population Prospects: The 2010 Revision. If you looked at population growth over a longer period of time, you would see that it is not actually linear. However, over the relatively short period of time above, the growth looks nearly linear. A statistical technique called linear regression can create a linear function that approximates the actual population growth over this period very well. It turns out that this function is P = 0.0024444t + 0.15914 where t represents the time variable measured in years since 1950 and P represents the (approximate) population of the U.S. measured in billions of people. (If you take statistics, you’ll probably learn how to obtain this function.) The graph of this linear function is shown in the figure above. (1) Just to make sure that you understand how to work with this function, use it to complete the following table. The actual population values are given. If you are working with the function correctly, the values you obtain should be close to the actual population values! Actual Population t P Year (billions of people) (years) (billions of people) 1960 .186158 1990 .256098 2005 .299846 (2) Use the linear function to determine the approximate year when the population of the U.S. -

Evolutionary Restoration Ecology
ch06 2/9/06 12:45 PM Page 113 189686 / Island Press / Falk Chapter 6 Evolutionary Restoration Ecology Craig A. Stockwell, Michael T. Kinnison, and Andrew P. Hendry Restoration Ecology and Evolutionary Process Restoration activities have increased dramatically in recent years, creating evolutionary chal- lenges and opportunities. Though restoration has favored a strong focus on the role of habi- tat, concerns surrounding the evolutionary ecology of populations are increasing. In this con- text, previous researchers have considered the importance of preserving extant diversity and maintaining future evolutionary potential (Montalvo et al. 1997; Lesica and Allendorf 1999), but they have usually ignored the prospect of ongoing evolution in real time. However, such contemporary evolution (changes occurring over one to a few hundred generations) appears to be relatively common in nature (Stockwell and Weeks 1999; Bone and Farres 2001; Kin- nison and Hendry 2001; Reznick and Ghalambor 2001; Ashley et al. 2003; Stockwell et al. 2003). Moreover, it is often associated with situations that may prevail in restoration projects, namely the presence of introduced populations and other anthropogenic disturbances (Stockwell and Weeks 1999; Bone and Farres 2001; Reznick and Ghalambor 2001) (Table 6.1). Any restoration program may thus entail consideration of evolution in the past, present, and future. Restoration efforts often involve dramatic and rapid shifts in habitat that may even lead to different ecological states (such as altered fire regimes) (Suding et al. 2003). Genetic variants that evolved within historically different evolutionary contexts (the past) may thus be pitted against novel and mismatched current conditions (the present). The degree of this mismatch should then determine the pattern and strength of selection acting on trait variation in such populations (Box 6.1; Figure 6.1). -

Utah's Long-Term Demographic and Economic Projections Summary Principal Researchers: Pamela S
Research Brief July 2017 Utah's Long-Term Demographic and Economic Projections Summary Principal Researchers: Pamela S. Perlich, Mike Hollingshaus, Emily R. Harris, Juliette Tennert & Michael T. Hogue continue the existing trend of a slow decline. From Background 2015-2065, rates are projected to decline from 2.32 to 2.29. These rates are projected to remain higher The Kem C. Gardner Policy Institute prepares long-term than national rates that move from 1.87 to 1.86 over demographic and economic projections to support in- a similar period. formed decision making in the state. The Utah Legislature funds this research, which is done in collaboration with • In 2065, life expectancy in Utah is projected to be the Governor’s Office of Management and Budget, the -Of 86.3 for women and 85.2 for men. This is an increase fice of the Legislative Fiscal Analyst, the Utah Association of approximately 4 years for women and 6 years for of Governments, and other research entities. These 50- men. The sharper increase for men narrows the life year projections indicate continued population growth expectancy gap traditionally seen between the and illuminate a range of future dynamics and structural sexes. shifts for Utah. An initial set of products is available online • Natural increase (births minus deaths) is projected to at gardner.utah.edu. Additional research briefs, fact remain positive and account for two-thirds of the cu- sheets, web-enabled visualizations, and other products mulative population increase to 2065. However, giv- will be produced in the coming year. en increased life expectancy and declining fertility, the rate and amount of natural increase are project- State-Level Results ed to slowly decline over time. -

Human Population 2018 Lecture 8 Ecological Footprint
Human Population 2018 Lecture 8 Ecological footprint. The Daly criterea. Questions from the reading. pp. 87-107 Herman Daly “All my economists say, ‘on the one hand...on the other'. Give me a one- handed economist,” demanded a frustrated Harry S Truman. BOOKS Daly, Herman E. (1991) [1977]. Steady-State Economics (2nd. ed.). Washington, DC: Island Press. Daly, Herman E.; Cobb, John B., Jr (1994) [1989]. For the Common Good: Redirecting the Economy toward Community, the Environment, and a Sustainable Future (2nd. updated and expanded ed.). Boston: Beacon Press.. Received the Grawemeyer Award for ideas for improving World Order. Daly, Herman E. (1996). Beyond Growth: The Economics of Sustainable Development. Boston: Beacon Press. ISBN 9780807047095. Prugh, Thomas; Costanza, Robert; Daly, Herman E. (2000). The Local Politics of Global Sustainability. Washington, DC: Island Press. IS The Daly Criterea for sustainability • For a renewable resource, the sustainable rate to use can be no more than the rate of regeneration of its source. • For a non-renewable resource, the sustainable rate of use can be no greater than the rate at which a renewable resource, used sustainably, can be substituted for it. • For a pollutant, the sustainable rate of emmission can be no greater that the rate it can be recycled, absorbed or rendered harmless in its sink. http://www.footprintnetwork.org/ Ecosystem services Herbivore numbers control Carbon capture and Plant oxygen recycling and production soil replenishment Soil maintenance and processing Carbon and water storage system Do we need wild species? (negative) • We depend mostly on domesticated species for food (chickens...). • Food for domesticated species is itself from domesticated species (grains..) • Domesticated plants only need water, nutrients and light. -

Population Biology & Life Tables
EXERCISE 3 Population Biology: Life Tables & Theoretical Populations The purpose of this lab is to introduce the basic principles of population biology and to allow you to manipulate and explore a few of the most common equations using some simple Mathcad© wooksheets. A good introduction of this subject can be found in a general biology text book such as Campbell (1996), while a more complete discussion of pop- ulation biology can be found in an ecology text (e.g., Begon et al. 1990) or in one of the references listed at the end of this exercise. Exercise Objectives: After you have completed this lab, you should be able to: 1. Give deÞnitions of the terms in bold type. 2. Estimate population size from capture-recapture data. 3. Compare the following sets of terms: semelparous vs. iteroparous life cycles, cohort vs. static life tables, Type I vs. II vs. III survivorship curves, density dependent vs. density independent population growth, discrete vs. continuous breeding seasons, divergent vs. dampening oscillation cycles, and time lag vs. generation time in population models. 4. Calculate lx, dx, qx, R0, Tc, and ex; and estimate r from life table data. 5. Choose the appropriate theoretical model for predicting growth of a given population. 6. Calculate population size at a particular time (Nt+1) when given its size one time unit previous (Nt) and the corre- sponding variables (e.g., r, K, T, and/or L) of the appropriate model. 7. Understand how r, K, T, and L affect population growth. Population Size A population is a localized group of individuals of the same species. -
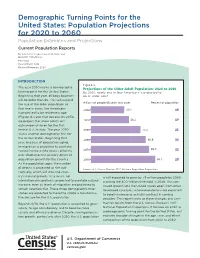
Population Projections for 2020 to 2060 Population Estimates and Projections Current Population Reports
Demographic Turning Points for the United States: Population Projections for 2020 to 2060 Population Estimates and Projections Current Population Reports By Jonathan Vespa, Lauren Medina, and David M. Armstrong P25-1144 Issued March 2018 Revised February 2020 INTRODUCTION Figure The year 2030 marks a demographic Projections of the Older Adult Population to turning point for the United States. By nearly one in four Americans is projected to Beginning that year, all baby boomers be an older adult will be older than 65. This will expand Millions of people years and older Percent of population the size of the older population so that one in every five Americans is projected to be retirement age (Figure 1). Later that decade, by 2034, we project that older adults will outnumber children for the first time in U.S. history. The year 2030 marks another demographic first for the United States. Beginning that year, because of population aging, immigration is projected to overtake natural increase (the excess of births over deaths) as the primary driver of population growth for the country. As the population ages, the number of deaths is projected to rise sub- Source US Census Bureau National Population Projections stantially, which will slow the coun- try’s natural growth. As a result, net is still expected to grow by 79 million people by 2060, international migration is projected to overtake natural crossing the 400-million threshold in 2058. This con- increase, even as levels of migration are projected to tinued growth sets the United States apart from other remain relatively flat. These three demographic mile- developed countries, whose populations are expected stones are expected to make the 2030s a transforma- to barely increase or actually contract in coming tive decade for the U.S. -

Global Population Trends: the Prospects for Stabilization
Global Population Trends The Prospects for Stabilization by Warren C. Robinson Fertility is declining worldwide. It now seems likely that global population will stabilize within the next century. But this outcome will depend on the choices couples make throughout the world, since humans now control their demo- graphic destiny. or the last several decades, world population growth Trends in Growth Fhas been a lively topic on the public agenda. For The United Nations Population Division makes vary- most of the seventies and eighties, a frankly neo- ing assumptions about mortality and fertility to arrive Malthusian “population bomb” view was in ascendan- at “high,” “medium,” and “low” estimates of future cy, predicting massive, unchecked increases in world world population figures. The U.N. “medium” variant population leading to economic and ecological catas- assumes mortality falling globally to life expectancies trophe. In recent years, a pronatalist “birth dearth” of 82.5 years for males and 87.5 for females between lobby has emerged, with predictions of sharp declines the years 2045–2050. in world population leading to totally different but This estimate assumes that modest mortality equally grave economic and social consequences. To declines will continue in the next few decades. By this divergence of opinion has recently been added an implication, food, water, and breathable air will not be emotionally charged debate on international migration. scarce and we will hold our own against new health The volatile mix has exploded into a torrent of threats. It further assumes that policymakers will books, scholarly articles, news stories, and op-ed continue to support medical, scientific, and technolog- pieces, presenting at least superficially plausible data ical advances, and that such policies will continue to and convincing arguments on all sides of every ques- have about the same effect on mortality as they have tion. -

Equilibrium Theory of Island Biogeography: a Review
Equilibrium Theory of Island Biogeography: A Review Angela D. Yu Simon A. Lei Abstract—The topography, climatic pattern, location, and origin of relationship, dispersal mechanisms and their response to islands generate unique patterns of species distribution. The equi- isolation, and species turnover. Additionally, conservation librium theory of island biogeography creates a general framework of oceanic and continental (habitat) islands is examined in in which the study of taxon distribution and broad island trends relation to minimum viable populations and areas, may be conducted. Critical components of the equilibrium theory metapopulation dynamics, and continental reserve design. include the species-area relationship, island-mainland relation- Finally, adverse anthropogenic impacts on island ecosys- ship, dispersal mechanisms, and species turnover. Because of the tems are investigated, including overexploitation of re- theoretical similarities between islands and fragmented mainland sources, habitat destruction, and introduction of exotic spe- landscapes, reserve conservation efforts have attempted to apply cies and diseases (biological invasions). Throughout this the theory of island biogeography to improve continental reserve article, theories of many researchers are re-introduced and designs, and to provide insight into metapopulation dynamics and utilized in an analytical manner. The objective of this article the SLOSS debate. However, due to extensive negative anthropo- is to review previously published data, and to reveal if any genic activities, overexploitation of resources, habitat destruction, classical and emergent theories may be brought into the as well as introduction of exotic species and associated foreign study of island biogeography and its relevance to mainland diseases (biological invasions), island conservation has recently ecosystem patterns. become a pressing issue itself.