Ecological Modelling 220 (2009) 2690–2701
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1 Anleitung Für Die Geographische Artendatenbank Nachdem Sie Die
Anleitung für die geographische Artendatenbank Nachdem Sie die Anwendung gestartet haben, können Sie mit den entsprechenden Werkzeugen zur gewünschten geographischen Lage finden. Im linken Auswahlmenü wählen Sie bitte "Artenfunde digitalisieren". Mit dem Button können Sie einen Punkt in die Karte setzen. Bitte beachten Sie unbedingt, dass bevor ein Punkt gesetzt wird alles geladen ist. Es müssen ungefähr 1,4 MB (Artenliste mit ca. 19.000 Arten) geladen werden. Links erscheint dann ein Disketten Symbol . Nach klick auf das Symbol erscheint ein Fenster, in dem die erforderlichen Angaben einzutragen sind. Die Felder bis „Ort des Fundes“ sind Pflichtfelder, hier müssen unbedingt Eingaben gemacht werden. 1 Die Eingabe über Autor und E-Mail des Autors sowie Bemerkungen sollten ebenso eingegeben werden. Diese Angaben werden in der Datenbank gespeichert, jedoch nicht veröffentlicht. Diese Angaben dienen intern dazu, die Wertigkeit der Eingaben beurteilen zu können. Es stehen z.B. beim "Artenname" Pulldown-Listen zur Verfügung, dadurch wird eine einheitliche Eingabe garantiert. Es stehen ca. 19.000 Arten zur Verfügung. Sollte es für eine Art keinen deutschen Namen geben, steht der wissenschaftliche Name zur Verfügung. Die Liste ist alphabetisch sortiert. Außerdem werden in der Liste keine ü,ö,ä und ß verwendet. Die Namen werden mit Umlauten geschrieben. Die vollständige Liste finden Sie im Anhang zu dieser Anleitung. Das Datum ist im Format JJJJ-MM-TT (z.B. 2012-01-27) einzugeben. Das wäre der 27. Januar 2012. Beenden Sie alle Eingaben durch drücken auf "Speichern". Während Ihrer aktuellen Internetsitzung haben Sie die Möglichkeit mit dem Button die Eingabe des Datensatzes wieder aus der Datenbank zu löschen. -

Botany Orary
TAXONOMY AND CULTIVAR DEVELOPMENT OF POA PRATENSIS L. David P. Byres 1 13 BOTANY ORARY PhD. University of Edinburgh 1984 1) Declaration. This thesis was composed by myself, and the work described herein is my own. David P. Byres CONTENTS Acknowledgements . • I \/ Abstract . Section A . Taxonomy. Chapter 1. Introduction. 1.1 Introduction ..........................................1 1.2 Taxonomy .............................................1 1.3 Cultivar Development ............................... 4 Chapter 2. Literature Review : Taxonomy 2.1 Introduction ............. .............................7 2.2 History of the Taxonomic Treatment of of Poa pratensis L. s.1............ ....... 10 2.3 Taxonomic Characters used by previous workers .........14 Chapter 3. Materials and Methods. 3.1 Environmental Variation .................... 20 3.2 Population and Herbarium Studies .......... ............26 3.3 Taxonomic Analysis of Biotypes and Cultivars... ... ... 36 3.4 Statistical Analysis ..................................37 Chapter 4. Effect of Environmental Variation on Morphology. 4.1 Introduction .......................................... 38 4.2 Results ........................................ .......38 Chapter 5. Study of Poa pratensis Populations. 5.1 Introduction .......................................... 49 5.2 Results ........................ 49 Chapter 6. Study of Herbarium Material. 6.1 Introduction ......................................... 60 6.2 Results ....................... 60 Chapter 7. Morphological Examination of Biotypes and -

Ecography ECOG-05013
Ecography ECOG-05013 Aikio, S., Ramula, S., Muola, A. and von Numers, M. 2020. Island properties dominate species traits in determining plant colonizations in an archipelago system. – Ecography doi: 10.1111/ecog.05013 Supplementary material Supplementary material Appendix 1 Fig. A1. Pairwise relationships and correlation coefficients of the island variables in the AIC- simplified model. 1 Fig. A2. Pairwise relationships and correlation coefficients of the plant traits in the AIC- simplified model. 2 Dispersal_vector [wind_water] Dispersal_vector [endozoochor] Dispersal_vector [unspecialised] Historical_total_log Pollen_vector [abiotic_insect] Life_form [herb] Dispersal_vector [myrmerochor] Dispersal_vector [epizoochor] Area_log Plant_height_log Limestone [Yes] Convolution Buffer_2_km_log Life_cycle [short] Seed_mass_log Veg_repr [Yes] North_limit Seed_bank [transient] Ellenberg_Nitrogen Ellenberg_Moisture Residents_per_area_log Ellenberg_Temperature Ellenberg_Reaction Shannon_habitats Shore_meadow Deciduous_forest Mixed_forest Marsh Buffer_5_km_log Buildings Euref_X Meadow_or_pasture Euref_Y Eklund_culture SLA Ellenberg_Light Coniferous_forest Sand Open_rock_or_bare_ground Seed_bank [unknown] Apomictic [Yes] Life_form [woody] Pollen_vector [insect] Pollen_vector [abiotic_self] Dist_to_historical_log Pollen_vector [self] Pollen_vector [insect_self] 0.2 0.5 1 2 5 10 Odds Ratio Fig. A3. Odds ratios (i.e., exp[parameter estimate]) and 95% confidence intervals for the fixed effects of the colonization model (full model of all 587 species with -

Flora of North America North of Mexico
Flora of North America North of Mexico Edited by FLORA OF NORTH AMERICA EDITORIAL COMMITTEE VOLUME 24 MagnoUophyta: Commelinidae (in part): Foaceae, part 1 Edited by Mary E. Barkworth, Kathleen M. Capéis, Sandy Long, Laurel K. Anderton, and Michael B. Piep Illustrated by Cindy Talbot Roché, Linda Ann Vorobik, Sandy Long, Annaliese Miller, Bee F Gunn, and Christine Roberts NEW YORK OXFORD • OXFORD UNIVERSITY PRESS » 2007 Oxford Univei;sLty Press, Inc., publishes works that further Oxford University's objective of excellence in research, scholarship, and education. Oxford New York /Auckland Cape Town Dar es Salaam Hong Kong Karachi Kuala Lumpur Madrid Melbourne Mexico City Nairobi New Delhi Shanghai Taipei Toronto Copyright ©2007 by Utah State University Tlie account of Avena is reproduced by permission of Bernard R. Baum for the Department of Agriculture and Agri-Food, Government of Canada, ©Minister of Public Works and Government Services, Canada, 2007. The accounts of Arctophila, Dtipontui, Scbizacbne, Vahlodea, xArctodiipontia, and xDiipoa are reproduced by permission of Jacques Cayouette and Stephen J. Darbyshire for the Department of Agriculture and Agri-Food, Government of Canada, ©Minister of Public Works and Government Services, Canada, 2007. The accounts of Eremopoa, Leitcopoa, Schedoiioms, and xPucciphippsia are reproduced by permission of Stephen J. Darbyshire for the Department of Agriculture and Agri-Food, Government of Canada, ©Minister of Public Works and Government Services, Canada, 2007. Published by Oxford University Press, Inc. 198 Madison Avenue, New York, New York 10016 www.oup.com Oxford is a registered trademark of Oxford University Press All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording, or otherwise, without the prior written permission of Utah State University. -

Diplomarbeit
Diplomarbeit vorgelegt zur Erlangung des Grades eines Diplom-Biologen an der Fakultät für Biologie und Biotechnologie der Ruhr-Universität Bochum Phylogenie der Brandpilzgattung Urocystis von Sascha Lotze-Engelhard Angefertigt im LS Evolution und Biodiversität der Pflanzen, AG Geobotanik Bochum im Dezember 2010 Referent: Prof. Dr. D. Begerow Koreferent: Prof. Dr. R. Tollrian 2 ҅Fungi have a profound impact on global ecosystems. They modify our habitats and are essential for many ecosystem functions. Fungi form soil, recycle nutrients, decay wood, enhance plant growth and cull plants from their environment. They feed us, poison us, parasitize us and cure us. They destroy our crops, homes and libraries, but they also produce valuable biochemicals, such as ethanol and antibiotics. For both practical and intellectual reasons it is important to provide a phylogeny of Fungi on which a classi- fication can be firmly based. ҆ (Blackwell u.a. 2006) Inhaltsverzeichnis 3 Inhaltsverzeichnis Inhaltsverzeichnis ........................................................................................................... 3 Abbildungsverzeichnis .................................................................................................... 6 Tabellenverzeichnis ........................................................................................................ 7 Abkürzungsverzeichnis .................................................................................................. 8 1 Einleitung ............................................................................................................ -

Collection of New Poa Genetic Diversity
Collection of new Poa genetic diversity Evelin WILLNER & Magdalena SEVCIKOVA IPK Gene Bank External Branch North, Malchow, Germany OSEVA PRO Ltd ., Grassland Research Station, ZbriZubri, Cz ech Rep u blic Evelin Willner - ”Collection of new genetic diversity of the genus Poa and documentation…” – IPK-Institutstag 2006, 10.10.2006 The EPDB – contents and applications European Poa database (()EPDB) http://poa.ipk-gatersleben.de Search for IPK Collection information Poa ex ploration Evelin Willner - ”Collection of new genetic diversity of the genus Poa and documentation…” – IPK-Institutstag 2006, 10.10.2006 European Poa Database Search form of the European Poa Database: select single field (first step) Evelin Willner - ”Collection of new genetic diversity of the genus Poa and documentation…” – IPK-Institutstag 2006, 10.10.2006 Search form of the European Poa Database: select single field (second step) European Poa Database Evelin Willner - ”Collection of new genetic diversity of the genus Poa and documentation…” – IPK-Institutstag 2006, 10.10.2006 European Poa Database Search form of the European Poa Database: select single field (third step) Evelin Willner - ”Collection of new genetic diversity of the genus Poa and documentation…” – IPK-Institutstag 2006, 10.10.2006 Collection of new Poa genetic diversity Project Title: Plant Exploration in Central Europe to Collect Poa Species for Crop Improvement Participants: Dr. Richard C. Johnson, USDA-ARS/Department of Crop and Soil Sciences, Washington State University Organizer Dr. David Huff, Associate -
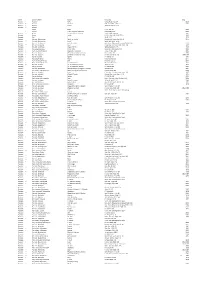
Tropicos Name Search Results Poa.Csv
Family ! Scientific Name Author Reference Date Poaceae * Poaceae Rchb. Consp. Regn. Veg. 54 1828 Poaceae !! Poaceae Barnhart Bull. Torrey Bot. Club 22: 7 1895 Poaceae * Poaceae Caruel Atti Reale Accad. Lincei Poaceae ** Poaeae Poales Small Fl. S.E. U.S. 48 1903 ! Poanae Takht. ex Reveal & Doweld Novon 9(4): 550 1999 Poaceae ** Poanae T.D. Macfarl. & L. Watson Taxon 31(2): 193–194 1982 Poaceae Poatae Hitchc. U.S.D.A. Bull. (1915‐23) 772: 6 1920 Poaceae Poa L. Sp. Pl. 1: 67 1753 Poaceae Poa sect. Abbreviatae Nannf. ex Tzvelev Novosti Sist. Vyssh. Rast. 11: 30 1974 Poaceae ** Poa sect. Abbreviatae Nannf. Symb. Bot. Upsal. 5: 25 Poaceae Poa sect. Acroleucae Prob. & Tzvelev Bot. Zhurn. [Moscow & Leningrad] 95(6): 864 2010 Poaceae ** Poa ser. Acroleucae Keng Claves Gen. Sp. Gram. Prim. Sinic. 169 1957 Poaceae Poa sect. Acutifoliae Pilg. ex Potztal Willdenowia 5(3): 473 1969 Poaceae Poa subg. Aeluropus (Trin.) Rchb. Deut. Bot. Herb.‐Buch 2: 39 1841 Poaceae ! Poa [ unranked ] Alpinae Hegetschw. ex Nyman Consp. Fl. Eur. 835 1882 Poaceae ** Poa sect. Alpinae (Hegetschw. ex Nyman) Soreng Novon 8(2): 193 1998 Poaceae ! Poa sect. Alpinae (Hegetschw. ex Nyman) Stapf Fl. Brit. India 7(22): 338 1897 [1896 Poaceae ! Poa ser. Alpinae Roshev. Fl. URSS 2: 411 1934 Poaceae Poa subg. Andinae Nicora Hickenia 1(18): 99 1977 Poaceae * Poa [ unranked ] Annuae Döll Fl. Baden 1: 172 1855 Poaceae ! Poa [ unranked ] Annuae Fr. ex Andersson Pl. Scand. Gram. 47 1852 Poaceae ** Poa sect. Annuae (Fr. ex Andersson) Lindm. Skand. Fl. 213 1926 Poaceae Poa ser. -
Bayer Expert Guide Meadow-Grass Management in Winter Cereals
Bayer Expert Guide Meadow-grass Management in Winter Cereals www.bayercropscience.co.uk Introduction Copyright of photographs The term ‘meadow-grasses’ refers broadly to a group of around fifteen Poa species found in the UK. A few species are widespread All photography © Bayer CropScience Ltd 2015, except where indicated. and common; others very rare with only three of significant importance to the farmer. These are annual meadow-grass (Poa annua), rough meadow-grass (Poa trivialis, also known Active substances and trademark acknowledgements as rough-stalked meadow-grass) and smooth meadow-grass (Poa pratensis, also known as smooth-stalked meadow-grass). Atlantis®, Bacara®, biopower®, DFF ®, Liberator®, Movon®, Othello® and Vigon® are registered trademarks of Bayer. Of the three, both annual meadow-grass and rough meadow-grass are commonly found as weeds in arable and horticultural crops Atlantis WG contains mesosulfuron-methyl and iodosulfuron-methyl-sodium. Bacara contains diflufenican and flurtamone. (1,9,38,41,43). Liberator contains flufenacet and diflufenican. Othello contains diflufenican, iodosulfuron-methyl-sodium and mesosulfuron-methyl. Movon and Vigon contain flufenacet, diflufenican and flurtamone. Although smooth meadow-grass is widespread The objective of this booklet is to aid and assist Use plant protection products safely. and frequently found as a constituent of pastures, in the identification of these more common Always read the label and product information before use. grassy banks, roadside verges and field margins, meadow-grasses, and give an insight into their Pay attention to the risk indications and follow the safety precautions on the label. it is not usually a serious or persistent problem distribution and biology, and guidance on their For further information, please visit www.bayercropscience.co.uk or call Bayer Assist weed in arable crops (1,41). -
Aberystwyth University the Reappearance of Lobelia Urens From
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Aberystwyth Research Portal Aberystwyth University The reappearance of Lobelia urens from soil seed bank at a site in South Devon Smith, R. E. N. Published in: Watsonia Publication date: 2002 Citation for published version (APA): Smith, R. E. N. (2002). The reappearance of Lobelia urens from soil seed bank at a site in South Devon. Watsonia, 107-112. http://hdl.handle.net/2160/4028 General rights Copyright and moral rights for the publications made accessible in the Aberystwyth Research Portal (the Institutional Repository) are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the Aberystwyth Research Portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the Aberystwyth Research Portal Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. tel: +44 1970 62 2400 email: [email protected] Download date: 09. Jul. 2020 Index to Watsonia vols. 1-25 (1949-2005) by Chris Boon Abbott, P. P., 1991, Rev. of Flora of the East Riding of Yorkshire (by E. Crackles with R. Arnett (ed.)), 18, 323-324 Abbott, P. -
Fjölrit Náttúrufræðistofnunar Annotated Checklist of Vascular Plants Iceland
FJÖLRIT NÁTTÚRUFRÆÐISTOFNUNAR ANNOTATED CHECKLIST OF VASCULAR PLANTS ICELAND ANNOTATED CHECKLIST OF VASCULAR PLANTS OF ICELAND Paweł Wąsowicz 57 57 FJÖLRIT NÁTTÚRUFRÆÐISTOFNUNAR ANNOTATED CHECKLIST OF VASCULAR PLANTS OF ICELAND Paweł Wąsowicz 57 FJÖLRIT NÁTTÚRUFRÆÐISTOFNUNAR Nr. 57, mars 2020 Fjölrit Náttúrufræðistofnunar er ritröð sem hóf göngu sína árið 1985. Birtar eru greinar og skýrslur eftir starfsmenn og fræðimenn sem vinna í samvinnu við þá. Í hverju hefti er ein sjálfstæð grein um náttúrufræði. Útgáfan er óregluleg. Greinar eru ritaðar á íslensku með enskum útdrætti. Þær mega einnig vera á ensku en þá skal ávallt fylgja ítarlegur útdráttur á íslensku. Vitnið til þessa rits á eftirfarandi hátt – Recommended citation: Paweł Wąsowicz 2020. Annotated checklist of vascular plants of Iceland. Fjölrit Náttúrufræðistofnunar nr. 57. Garðabær: Náttúrufræðistofnun Íslands. DOI: 10.33112/1027-832X.57 Ritnefnd María Harðardóttir, Guðmundur Guðmundsson og Guðríður Gyða Eyjólfsdóttir Kápumynd Holtasóley (Dryas octopetala) Stílfærð teikning Anette Theresia Meier eftir teikningu Carl Axel Magnus Lindman úr bókinni Bilder ur Nordens Flora Umbrot María Harðardóttir Útgefandi NÁTTÚRUFRÆÐISTOFNUN ÍSLANDS Urriðaholtsstræti 6–8 210 Garðabæ Sími: 590 0500 Netfang: [email protected] www.ni.is Prentun Prentmet Oddi ©Náttúrufræðistofnun Íslands 2020 ISSN 1027-832X ANNOTATED CHECKLIST OF VASCULAR PLANTS OF ICELAND Paweł Wąsowicz EFNISYFIRLIT ABSTRACT 5 INTRODUCTION 7 DEFINITIONS OF TERMS AND ABBREVIATIONS USED 10 CHECKLIST OF VASCULAR PLANTS – PLÖNTUTAL 11 ÚTDRÁTTUR Á ÍSLENSKU 167 REFERENCES – HEIMILDIR 169 INDEX OF GENERA – SKRÁ YFIR LATNESK ÆTTKVÍSLAHEITI 174 INDEX OF ICELANDIC NAMES – SKRÁ YFIR TEGUNDAHEITI 180 FJÖLRIT NÁTTÚRUFRÆÐISTOFNUNAR 192 – 3 – FJÖLRIT 57 NÁTTÚRUFRÆÐISTOFNUN ÍSLANDS, mars 2020 ANNOTATED CHECKLIST OF VASCULAR PLANTS OF ICELAND Paweł Wąsowicz ABSTRACT The present edition of the annotated checklist is a non-native of unknown age and 19 taxa qualified as comprehensive catalogue of all vascular plant taxa: archaeophytes. -

Diversity of Plant Species Eaten and Dispersed by the European Bison Bison Bonasus in Białowieża Forest
European Bison Conservation Newsletter Vol 1 (2008) pp: 14–29 Diversity of plant species eaten and dispersed by the European bison Bison bonasus in Białowieża Forest Bogdan Jaroszewicz1, Ewa Pirożnikow2 1 Białowieża Geobotanical Station, Institute of Botany at the Faculty of Biology, University of Warsaw, Sportowa 19, 17–230 Białowieża; e-mail: [email protected] 2 Institute of Biology, University of Białystok Abstract The European bison influences vegetation by e.g. endozoochoric seed dispersal. The diet of the European bison includes 454 vascular plant species, i.e. close to 40% of the flora of Białowiez˙a Forest. Seeds contained in 75 samples of bison faeces developed into 12,392 seedlings, classified as 173 different plant species, of which 80% were perennials. Species whose seeds are disseminated via the gastrointestinal tract of the bison constitute 16% of the vascular plants recorded in Białowiez˙a Forest. The total number of plant species dispersed by bison is close to double the number recorded for domestic cattle, horses or Cervidae. The biodiversity of plant species dispersed by free-roaming bison is lower (102 species) than that of bison living in captivity (140 species). In the faeces of both free-roaming and captive bison the number of anthropogenic and meadow plant species was higher in winter than in summer. The European bison plays a significant role in the dispersal of plants, especially of non-forest species, which ensures their existence in the forest community. Plant seeds present in bison faeces which do not naturally occur in Białowiez˙a Forest (probably introduced with hay used for supplementary feeding) contributes to the stability of flora biodiversity, but could also introduce the risk of invasion by alien species. -

Remote Sensing and GIS for Wetland Vegetation Study
Al Sghair, Fathi Goma (2013) Remote sensing and GIS for wetland vegetation study. PhD thesis http://theses.gla.ac.uk/4581/ Copyright and moral rights for this thesis are retained by the author A copy can be downloaded for personal non-commercial research or study, without prior permission or charge This thesis cannot be reproduced or quoted extensively from without first obtaining permission in writing from the Author The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the Author When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given. Glasgow Theses Service http://theses.gla.ac.uk/ [email protected] Remote Sensing and GIS for Wetland Vegetation Study Fathi Goma Al Sghair This thesis is submitted in fulfilment of the requirements for the Degree of Doctor of Philosophy Institute of Biodiversity, Animal Health and Comparative Medicine College of Medical, Veterinary and Life Sciences University of Glasgow September 2013 ii Abstract Remote Sensing (RS) and Geographic Information System (GIS) approaches, combined with ground truthing, are providing new tools for advanced ecosystem management, by providing the ability to monitor change over time at local, regional, and global scales. In this study, remote sensing (Landsat TM and aerial photographs) and GIS, combined with ground truthing work, were used to assess wetland vegetation change over time at two contrasting wetland sites in the UK: freshwater wetland at Wicken Fen between 1984 and 2009, and saltmarsh between 1988 and 2009 in Caerlaverock Reserve.