Ophiothrix Trindadensis Tommasi, 1970
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Life History Patterns of Three Estuarine Brittlestars (Ophiuroidea) at Cedar Key, Florida
LIFE HISTORY PATTERNS OF THREE ESTUARINE BRITTLESTARS (OPHIUROIDEA) AT CEDAR KEY, FLORIDA by STEPHEN EDWARD STANCYK A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 1974 UNIVERSITY OF FLORIDA 3 1262 08552 5847 ^ ACKNOWLEDGMENTS I owe a great" deal to all the people who made the completion of this dissertation possible. The members of my committee deserve special tnanks, particularly Drs. Frank Maturo and Thomas Emmel , who were always ready with encouragement and advice. I thank Drs. John Erookbank and Ariel Lugo for their careful reading of the manuscript, and Dr. John Ewel for his timely services. Dr. John Anderson was generous with both equipment and time. Of the many fellow students and friends who assisted me, William Ingram deserves special thanks for his indispensable aid in fostering an agreeable relationship between myself and the computer. John Caldwell, John Paige, Christine Simon and Michael Oesterling were of particular help in the field, and I would like to thank Dave David, Renee Lindsay,- Kent Murphey, Dave Godman and Steve Salzman for their assistance in the. laboratory. Marine biologists are often in need of a sa c e haven in a storm, and V.am therefore very grateful to Lee and Esta Belcher and thei wonderful family for their hospitality, and for making my work at Cedar Key such a pleasurable experience. Ms-:. Lib by Coker typed the final manuscript, and Mr. Paul Laessle provided. materia Is and advice for completion of the figures. -

Jacksonville, Florida 1998 Odmds Benthic Community Assessment
JACKSONVILLE, FLORIDA 1998 ODMDS BENTHIC COMMUNITY ASSESSMENT Submitted to U.S. Environmental Protection Agency, Region 4 61 Forsyth St. Atlanta, Georgia 30303 Prepared by Barry A. Vittor & Associates, Inc. 8060 Cottage Hill Rd. Mobile, Alabama 36695 (334) 633-6100 November 1999 TABLE OF CONTENTS LIST OF TABLES ………………………………………….……………………………3 LIST OF FIGURES ……………………..………………………………………………..4 1.0 INTRODUCTION ………..…………………………………………………………..5 2.0 METHODS ………..…………………………………………………………………..5 2.1 Sample Collection And Handling ………………………………………………5 2.2 Macroinfaunal Sample Analysis ……………………………………………….6 3.0 DATA ANALYSIS METHODS ……..………………………………………………6 3.1 Assemblage Analyses ..…………………………………………………………6 3.2 Faunal Similarities ……………………………………………………….…….8 4.0 HABITAT CHARACTERISTICS ……………………………………………….…8 5.0 BENTHIC COMMUNITY CHARACTERIZATION ……………………………..9 5.1 Faunal Composition, Abundance, And Community Structure …………………9 5.2 Numerical Classification Analysis …………………………………………….10 5.3 Taxa Assemblages …………………………………………………………….11 6.0 1995 vs 1998 COMPARISONS ……………………………………………………..11 7.0 SUMMARY ………………………………………………………………………….13 8.0 LITERATURE CITED ……………………………………………………………..16 2 LIST OF TABLES Table 1. Station locations for the Jacksonville, Florida ODMDS, June 1998. Table 2. Sediment data for the Jacksonville, Florida ODMDS, June 1998. Table 3. Summary of abundance of major taxonomic groups for the Jacksonville, Florida ODMDS, June 1998. Table 4. Abundance and distribution of major taxonomic groups at each station for the Jacksonville, Florida ODMDS, June 1998. Table 5. Abundance and distribution of taxa for the Jacksonville, Florida ODMDS, June 1998. Table 6. Percent abundance of dominant taxa (> 5% of the total assemblage) for the Jacksonville, Florida ODMDS, June 1998. Table 7. Summary of assemblage parameters for the Jacksonville, Florida ODMDS stations, June 1998. Table 8. Analysis of variance table for density differences between stations for the Jacksonville, Florida ODMDS stations, June 1998. -

Tube Epifaum of the Polychaete Phyllopchaetopterus Socialis
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Repository Open Access to Scientific Information from Embrapa Estuarine, Coastal and Shelf Science (1995) 41, 91–100 Tube epifauna of the Polychaete Phyllochaetopterus socialis Claparède Rosebel Cunha Nalessoa, Luíz Francisco L. Duarteb, Ivo Pierozzi Jrc and Eloisa Fiorim Enumod aDepartamento de Zoologia, CCB, Universidade Federal de Pernambuco, 50670-901, Recife, PE, Brazil, bDepartamento de Zoologia, Instituto Biologia, C.P. 6109, Universidade Estadual de Campinas, 13.081-970, Campinas, SP, Brazil, cEmbrapa, NMA, Av. Dr. Julio Soares de Arruda, 803 CEP 13.085, Campinas, SP, Brazil and dProtebras, Rua Turmalina, 79 CEP 13.088, Campinas, SP, Brazil Received 8 October 1992 and in revised form 22 June 1994 Keywords: Polychaeta; tubes; faunal association; epifauna; São Sebastião Channel; Brazil Animals greater than 1 mm, found among tangled tubes of Phyllochaetopterus socialis (Chaetopteridae) from Araçá Beach, São Sebastião district, Brazil, were studied for 1 year, with four samples in each of four seasons. They comprised 10 338 individuals in 1722·7 g dry weight of polychaete tubes, with Echino- dermata, Polychaeta (not identified to species) and Crustacea as the dominant taxa. The Shannon–Wiener diversity index did not vary seasonally, only two species (a holothurian and a pycnogonid) showing seasonal variation. Ophiactis savignyi was the dominant species, providing 45·5% of individuals. Three other ophiuroids, the holothurian Synaptula hidriformis, the crustaceans Leptochelia savignyi, Megalobrachium soriatum and Synalpheus fritzmuelleri, the sipunculan Themiste alutacea and the bivalve Hiatella arctica were all abundant, but most of the 68 species recorded occurred sparsely. -

UNIVERSITY of KERALA Zoology Core Course
1 UNIVERSITY OF KERALA First Degree Programme in Zoology Choice Based Credit and Semester System Zoology Core Course Syllabus-2015 Admission Onwards 2 FIRST DEGREE PROGRAMME IN ZOOLOGY Scheme of Instruction and Evaluation Course Study Components Instructional Credit Duration Evaluation Total Code Hrs/week of Univ. Credit T P Exam CE ESE Semster EN1111 English I 5 4 3 Hrs 20% 80% 1111 Additional language I 4 3 3 Hrs 20% 80% EN 1121 Foundation course I 4 2 3 Hrs 20% 80% CH1131.4 Complementary course I 2 2 3 Hrs 20% 80% Complementary course I 2 16 I Practical of CH1131.4 BO1131 Complementary course II 2 2 3 Hrs 20% 80% Complementary course II 2 Practical of BO1131 ZO1141 Core Course I 3 3 3 Hrs 20% 80% Core Course Practical of ZO1141 1 EN1211 English II 4 3 3 Hrs 20% 80% EN1212 English III 5 4 3 Hrs 20% 80% 1211 Additional language II 4 3 3 Hrs 20% 80% CH1231.4 Complementary course III 2 2 3 Hrs 20% 80% II Complementary course III 2 Practical of CH1231.4 17 BO1231 Complementary course IV 2 2 3 Hrs 20% 80% Complementary course II 2 Practical of BO1231 ZO1241 Core Course II 3 3 3 Hrs 20% 80% Core Course Practical of ZO1241 1 III EN1311 English IV 5 4 3 Hrs 20% 80% EN1312 Additional language III 5 4 3 Hrs 20% 80% CH1331 Complementary course V 3 3 3 Hrs 20% 80% CH1331.4 Complementary course V 2 Practical of CH1331.4 BO1331 Complementary course VI 3 3 3 Hrs 20% 80% 17 BO1332 Complementary course VI 2 Practical of BO1331 ZO1341 Core Course III 3 3 3 Hrs 20% 80% ZO1341 Core Course Practical of ZO1341 2 IV EN1411 English V 5 4 3 Hrs 20% 80% EN1411 Additional language II 5 4 3 Hrs 20% 80% CH1431.4 Complementary course VII 3 3 3 Hrs 20% 80% CH1432.4 Complementary course 2 4 3 Hrs 20% 80% Practical of CH1131.4, CH1231.4, CH1331.4, CH1431.4. -

Key to the Common Shallow-Water Brittle Stars (Echinodermata: Ophiuroidea) of the Gulf of Mexico and Caribbean Sea
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/228496999 Key to the common shallow-water brittle stars (Echinodermata: Ophiuroidea) of the Gulf of Mexico and Caribbean Sea Article · January 2007 CITATIONS READS 10 702 1 author: Christopher Pomory University of West Florida 34 PUBLICATIONS 303 CITATIONS SEE PROFILE All content following this page was uploaded by Christopher Pomory on 21 May 2014. The user has requested enhancement of the downloaded file. All in-text references underlined in blue are added to the original document and are linked to publications on ResearchGate, letting you access and read them immediately. 1 Key to the common shallow-water brittle stars (Echinodermata: Ophiuroidea) of the Gulf of Mexico and Caribbean Sea CHRISTOPHER M. POMORY 2007 Department of Biology, University of West Florida, 11000 University Parkway, Pensacola, FL 32514, USA. [email protected] ABSTRACT A key is given for 85 species of ophiuroids from the Gulf of Mexico and Caribbean Sea covering a depth range from the intertidal down to 30 m. Figures highlighting important anatomical features associated with couplets in the key are provided. 2 INTRODUCTION The Caribbean region is one of the major coral reef zoogeographic provinces and a region of intensive human use of marine resources for tourism and fisheries (Aide and Grau, 2004). With the world-wide decline of coral reefs, and deterioration of shallow-water marine habitats in general, ecological and biodiversity studies have become more important than ever before (Bellwood et al., 2004). Ecological and biodiversity studies require identification of collected specimens, often by biologists not specializing in taxonomy, and therefore identification guides easily accessible to a diversity of biologists are necessary. -

Two New Brittle Star Species of the Genus Ophiothrix
Caribbean Journal of Science, Vol. 41, No. 3, 583-599, 2005 Copyright 2005 College of Arts and Sciences University of Puerto Rico, Mayagu¨ez Two New Brittle Star Species of the Genus Ophiothrix (Echinodermata: Ophiuroidea: Ophiotrichidae) from Coral Reefs in the Southern Caribbean Sea, with Notes on Their Biology GORDON HENDLER Natural History Museum of Los Angeles County, 900 Exposition Boulevard, Los Angeles, California 90007, U.S.A. [email protected] ABSTRACT.—Two new species, Ophiothrix stri and Ophiothrix cimar, inhabit shallow reef-platforms and slopes in the Southern Caribbean, and occur together at localities in Costa Rica and Panama, nearly to Colombia. What appears to be an undescribed species resembling O. cimar has been reported from eastern Venezuela. In recent years, reefs where the species were previously observed have deteriorated because of environmental degradation. As a consequence, populations of the new species may have been reduced or eradicated. The new species have previously been mistaken for O. angulata, O. brachyactis, and O. lineata. Ophiothrix lineata, O. stri, and O. cimar have in common a suite of morphological features pointing to their systematic affinity, and a similar pigmentation pattern consisting of a thin, dark, medial arm stripe flanked by two pale stripes. Ophiothrix lineata is similar to Indo-Pacific members of the subgenus Placophiothrix and closely resembles Ophiothrix stri. The latter is extremely similar to O. synoecina, from Colombia, and both can live in association with the rock-boring echinoid Echinometra lucunter. Although O. synoecina is a protandric hermaphrodite that reportedly broods its young externally, the new species are gonochoric and do not brood. -

The Reproductive System and Associated Organs of the Brittle-Star Ophiothrix Fragilis
The Reproductive System and associated organs of the Brittle-star Ophiothrix fragilis. By J. B. Smith, M.A., PhJ)., Zoological Laboratory, Cambridge. With 15 Text-figures. CONTENTS. PAGE IHTKODTTCTION 267 METHODS 268 THE A-itTAT. OEGAN AXD BELATED SINUSES 269 The Left Axial Sinus and Organ ...... 279 The Right Axial Sinus and Organ ...... 280 THE GENITAL RACHIS 281 THE GONADS 287 Testis 288 Ovary 289 THE GoNODtrcrs 297 THE GENITAL BTTBSAE ........ 301 CoiTCLTrsiONs .......... 305 STTMMAEY 307 REFERENCES .......... 308 INTEODUCTION. The observations which are recorded in this paper are the outcome of an investigation directed, primarily, towards the interpretation of the morphology and histology of the nervous system of the ophiuroid Opldothrix fragilis, Abildgaard. During the course of the work opportunity was afforded for examination of large numbers of brittle-stars ranging from post- metamorphic individuals to fully mature specimens, the sizes being of a disc diameter of 0-4 to 16 mm. Prom observations made on living animals and by the examination of sectioned material it has been possible to obtain information relating to the morphology and development of the organs assoeiatedj either directly or indirectly, -with reproduction. The systems of organs described include the gonads, the gonodncts, the axial organ and related sinuses, the genital rachis and the 268 J. E. SMITH genital bursae. Some explanation of the inclusion of the axial organ complex, which is not primarily associated with the reproductive system, in this account is, perhaps, due. Two reasons may be given. The first is that the axial organ and the genital rachis, though of different origin, are in close association during development and throughout adult life, and the second is that the interpretation of the morphology of the axial organ complex, here given, is in close agreement with that of Fedotov (1924) who, in ascribing a double origin and structure to the axial organ and sinus system of Opbiuroids, is in disagreement with all earlier investigators of the problem. -

New Echinoderm Remains in the Buried Offerings of the Templo Mayor of Tenochtitlan, Mexico City
New echinoderm remains in the buried offerings of the Templo Mayor of Tenochtitlan, Mexico City Carolina Martín-Cao-Romero1, Francisco Alonso Solís-Marín2, Andrea Alejandra Caballero-Ochoa4, Yoalli Quetzalli Hernández-Díaz1, Leonardo López Luján3 & Belem Zúñiga-Arellano3 1. Posgrado en Ciencias del Mar y Limnología, UNAM, México; [email protected], [email protected] 2. Laboratorio de Sistemática y Ecología de Equinodermos, Instituto de Ciencias del Mar y Limnología (ICML), Universidad Nacional Autónoma de México, México; [email protected] 3. Proyecto Templo Mayor (PTM), Instituto Nacional de Antropología e Historia, México (INAH). 4. Facultad de Ciencias, Universidad Nacional Autónoma de México (UNAM), Circuito Exterior s/n, Ciudad Universitaria, Apdo. 70-305, Ciudad de México, México, C.P. 04510; [email protected] Received 01-XII-2016. Corrected 02-V-2017. Accepted 07-VI-2017. Abstract: Between 1978 and 1982 the ruins of the Templo Mayor of Tenochtitlan were exhumed a few meters northward from the central plaza (Zócalo) of Mexico City. The temple was the center of the Mexica’s ritual life and one of the most famous ceremonial buildings of its time (15th and 16th centuries). More than 200 offerings have been recovered in the temple and surrounding buildings. We identified vestiges of 14 species of echino- derms (mostly as disarticulated plates). These include six species of sea stars (Luidia superba, Astropecten regalis, Astropecten duplicatus, Phataria unifascialis, Nidorellia armata, Pentaceraster cumingi), one ophiu- roid species (Ophiothrix rudis), two species of sea urchins (Eucidaris thouarsii, Echinometra vanbrunti), four species of sand dollars (Mellita quinquiesperforata, Mellita notabilis, Encope laevis, Clypeaster speciosus) and one species of sea biscuit (Meoma ventricosa grandis). -

Echinoderm Species of Flower Garden Banks National Marine Sanctuary
CORAL CAP SPECIES OF FLOWER GARDEN BANKS NATIONAL MARINE SANCTUARY Classification Common name Scientific Name Echinoderms Brittle Stars Amphiodia pulchella Amphioplus tumidus Asteroschema sp. Astrocyclus caecilia Basket Star Astrophyton muricatum Asteroporpa annulata Gorgonocephalus sp. Ophiactis algicola Five-armed Ophiactis Ophiactis quinqueradia Savigny's Brittle Star Ophiactis savignyi Blunt-spined/Spiny Brittle Star Ophiocoma echinata Red Ophiocoma Ophiocoma wendtii Ophiocomella ophiactoides Banded-arm/Harlequin Brittle Star Ophioderma appressum Serpent Star Ophioderma brevispinum Ruby Brittle Star Ophioderma rubicundum Scaly Brittle Star, Red Serpent Star Ophioderma squamosissimum Elegant Brittle Star Ophiolepis elegans ?Ophiophragmus sp. Reticulated Brittle Star Ophionereis reticulata Ophiostigma isacanthus Angular Brittle Star Ophiothrix angulata Suenson's/Sponge Brittle Star Ophiothrix suensonii Ophiurochaeta littoralis Sea Cucumbers Five-toothed Sea Cucumber Actinopyga agassizii Beaded Sea Cucumber Euapta lappa Holothuria lentiginosa Chocolate Chip/Three-rowed Cucumber Isostichopus (Stichopus) badionotus Cucumber Thyone pseudofusus Sea Stars Astropecten comptus Asterinopsis lymani Asterinopsis pilosis Coronaster briareus Goniaster tessellatus Common Comet Star Linckia guildingii Linckia nodosa Guilding's/Comet Star Ophidiaster guildingii Sclerasterias contorta CORAL CAP SPECIES OF FLOWER GARDEN BANKS NATIONAL MARINE SANCTUARY Classification Common name Scientific Name Sea Urchins Purple-spined Sea Urchin Arbacia punctulata Centrostephanus longispinus rubricingulus Coelopleurus floridanus Long-spined Sea Urchin Diadema antillarum Slate-pencil Urchin Eucidaris tribuloides Genocidaris maculata Lytechinus euerces Green/Variegated Urchin Lytechinus variegatus Red Heart Urchin Meoma ventricosa Paraster doederleini Stained Collector Urchin Pseudoboletia maculata maculata Red Sea Urchin Stylocidaris affinis . -

Echinodermata, Ophiuroidea)
Vol. 16: 105–113, 2012 AQUATIC BIOLOGY Published online July 19 doi: 10.3354/ab00435 Aquat Biol Slow arm regeneration in the Antarctic brittle star Ophiura crassa (Echinodermata, Ophiuroidea) Melody S. Clark*, Terri Souster British Antarctic Survey, Natural Environment Research Council, High Cross, Madingley Road, Cambridge CB3 0ET, UK ABSTRACT: Regeneration of arms in brittle stars is thought to proceed slowly in low temperature environments. Here a survey of natural arm damage and arm regeneration rates is documented in the Antarctic brittle star Ophiura crassa. This relatively small ophiuroid, a detritivore found amongst red macroalgae, displays high levels of natural arm damage and repair. This is largely thought to be due to ice damage in the shallow waters it inhabits. The time scale of arm regener- ation was measured in an aquarium-based 10 mo experiment. There was a delayed regeneration phase of 7 mo before arm growth was detectable in this species. This is 2 mo longer than the longest time previously described, which was in another Antarctic ophiuroid, Ophionotus victo- riae. The subsequent regeneration of arms in O. crassa occurred at a rate of approximately 0.16 mm mo−1. To date, this is the slowest regeneration rate known of any ophiuroid. The confir- mation that such a long delay before arm regeneration occurs in a second Antarctic species pro- vides strong evidence that this phenomenon is yet another characteristic feature of Southern Ocean species, along with deferred maturity, slowed growth and development rates. It is unclear whether delayed initial regeneration phases are adaptations to, or limitations of, low temperature environments. -

Living Resources Report Texas A&M University-Corpus Christi Results - Open Bay Habitat
Center for Coastal Studies CCBNEP Living Resources Report Texas A&M University-Corpus Christi Results - Open Bay Habitat B. Living Resources - Habitats Detailed community profiles of estuarine habitats within the CCBNEP study area are not available. Therefore, in the following sections, the organisms, community structure, and ecosystem processes and functions of the major estuarine habitats (Open Bay, Oyster Reef, Hard Substrate, Seagrass Meadow, Coastal Marsh, Tidal Flat, Barrier Island, and Gulf Beach) within the CCBNEP study area are presented. The following major subjects will be addressed for each habitat: (1) Physical setting and processes; (2) Producers and Decomposers; (3) Consumers; (4) Community structure and zonation; and (5) Ecosystem processes. HABITAT 1: OPEN BAY Table Of Contents Page 1.1. Physical Setting & Processes ............................................................................ 45 1.1.1 Distribution within Project Area ......................................................... 45 1.1.2 Historical Development ....................................................................... 45 1.1.3 Physiography ...................................................................................... 45 1.1.4 Geology and Soils ................................................................................ 46 1.1.5 Hydrology and Chemistry ................................................................... 47 1.1.5.1 Tides .................................................................................... 47 1.1.5.2 Freshwater -
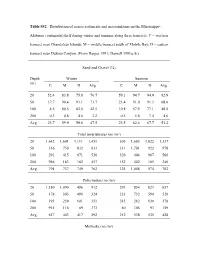
Table S32. Distribution of Coarse Sediments and Macroinfauna on the Mississippi
Table S32. Distribution of coarse sediments and macroinfauna on the Mississippi- Alabama continental shelf during winter and summer along three transects. C = western transect near Chandeleur Islands; M = middle transect south of Mobile Bay; D = eastern transect near DeSoto Canyon. (From Harper 1991; Darnell 1991a, b.) Sand and Gravel (%) Depth Winter Summer (m) C M D Avg. C M D Avg. 20 52.4 83.8 75.8 70.7 59.1 94.7 94.9 82.9 50 37.7 90.4 93.1 73.7 23.4 91.0 91.3 68.6 100 4.5 60.5 62.6 42.5 10.8 57.9 77.1 48.6 200 0.3 4.8 4.6 3.2 0.5 5.8 7.4 4.6 Avg. 23.7 59.9 59.0 47.5 23.5 62.4 67.7 51.2 Total invertebrates (no./m²) 20 1,642 1,601 1,111 1,451 505 1,683 1,822 1,337 50 316 750 832 633 311 1,701 922 978 100 291 415 871 526 326 404 967 566 200 946 183 182 457 152 402 185 246 Avg. 794 737 749 762 324 1,048 974 782 Polychaetes (no./m²) 20 1,180 1,090 486 912 293 854 823 657 50 178 305 490 324 233 732 594 520 100 193 259 601 351 243 262 630 378 200 915 116 89 373 80 305 93 159 Avg. 617 443 417 492 212 538 535 428 Mollusks (no./m²) 20 123 127 174 141 89 527 407 341 50 46 214 115 125 11 653 73 246 100 32 80 23 45 20 63 38 40 200 14 7 10 10 26 27 9 21 Avg.