Theories of Infant Development Theories of Infant Development
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Friends, Enemies, and Fools: a Collection of Uyghur Proverbs by Michael Fiddler, Zhejiang Normal University
GIALens 2017 Volume 11, No. 3 Friends, enemies, and fools: A collection of Uyghur proverbs by Michael Fiddler, Zhejiang Normal University Erning körki saqal, sözning körki maqal. A beard is the beauty of a man; proverbs are the beauty of speech. Abstract: This article presents two groups of Uyghur proverbs on the topics of friendship and wisdom, to my knowledge only the second set of Uyghur proverbs published with English translation (after Mahmut & Smith-Finley 2016). It begins with a brief introduction of Uyghur language and culture, then a description of Uyghur proverb styles, then the two sets of proverbs, and finally a few concluding comments. Key words: Proverb, Uyghur, folly, wisdom, friend, enemy 1. Introduction Uyghur is spoken by about 10 million people, mostly in northwest China’s Xinjiang Uyghur Autonomous Region. It is a Turkic language, most closely related to Uzbek and Salar, but also sharing similarity with its neighbors Kazakh and Kyrgyz. The lexicon is composed of about 50% old Turkic roots, 40% relatively old borrowings from Arabic and Persian, and 6% from Russian and 2% from Mandarin (Nadzhip 1971; since then, the Russian and Mandarin borrowings have presumably seen a slight increase due to increased language contact, especially with Mandarin). The grammar is generally head-final, with default SOV clause structure, A-N noun phrases, and postpositions. Stress is typically word-final. Its phoneme inventory includes eight vowels and 24 consonants. The phonology includes vowel harmony in both roots and suffixes, which is sensitive to rounded/unrounded (labial harmony) and front/back (palatal harmony), consonant harmony across suffixes, which is sensitive to voiced/unvoiced and front/back distinctions; and frequent devoicing of high vowels (see Comrie 1997 and Hahn 1991 for details). -

A Review of Heat Stroke and Its Complications in the Canine Renee L
Volume 45 | Issue 1 Article 1 1983 A Review of Heat Stroke and Its Complications in the Canine Renee L. Larson Iowa State University Robert W. Carithers Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/iowastate_veterinarian Part of the Small or Companion Animal Medicine Commons, and the Veterinary Physiology Commons Recommended Citation Larson, Renee L. and Carithers, Robert W. (1983) "A Review of Heat Stroke and Its Complications in the Canine," Iowa State University Veterinarian: Vol. 45 : Iss. 1 , Article 1. Available at: https://lib.dr.iastate.edu/iowastate_veterinarian/vol45/iss1/1 This Article is brought to you for free and open access by the Journals at Iowa State University Digital Repository. It has been accepted for inclusion in Iowa State University Veterinarian by an authorized editor of Iowa State University Digital Repository. For more information, please contact [email protected]. A Review of Heat Stroke and Its Complications in the Canine Renee L. Larson, BS * Robert W. Carithers, DVM, MS, PhD** SUMMARY most obvious prerequisite is a high en A review of heat stroke and its complications vironmental temperature. When the ambient is presented. The etiology, physiology, clinical temperature increases above 86°F, a rise in in signs, secondary complications, diagnosis, ternal body temperature results. Dogs can treatment, necropsy results and prevention of tolerate rising environmental temperature heat stroke are discussed. A clinical case is quite well. However, when the body tempera then presented to illustrate the disorder of heat ture exceeds 104°F a breakdown of the stroke. animal's thermal equilibrium begins. -

T'ahsíi Gha Gots'ęh Náets'ehndı́h Gha Goɂǫ
CANADA GOGHA AMÍI GOHÉH DZÁA NĮONIDHE SÍI MEGHA ?E?Á T’AHSÍI GHA GOTS’ĘH NÁETS’EHNDÍH GHA GOɁǪ agots’ı̨ la t’áh ǫkı edáidzę gots’ę́ gots’áendíh íle gha T’ahsíi gha gots’ęh naets’éhndı́h énidé gots’áendıh́ ts’ęh edı̨ htł’éh gogháts’ı̨ ɂa gots’ęh gha go ǫ azhíi ǫt’e? eghálats’enda íle gha godı̨ eghálats’enda éhsíi edı̨ htł’éh ɂ seets’eleh gogháts’ıdhah.́ T’ahsíi goghǫ ts’enéhe T’ahsíi gha gots’ęh náets’éhndıh́ gha goɂǫ énidé t’áh dádéhtí íle énidé gots’ęh t’ahsíi tsıts’ı́ ̨ hthe dádéhtí amíi méh dzaah nıgot’ǫ́ ts’ıhɂǫh gosaamba hule agújá, náts’ehndíh mets’ęh edı̨ htł’éh ats’ehɂı̨ gogháts’ı̨ ɂá íle amíi mets’ę́ t’áhsíi ts’enıdhę t’áh. T’ahsíi gha gots’ęh énidé dádétí náíndí ts’enidhę t’áh gots’ęnoendíh. náets’éhndıh́ gha goɂǫ énidé t’áh amíi t’ahsíi tsı̨ guıdhe meghǫh názhaetı gots’eh zǫ́ dúle gots’ęh noendıh́ Gohseenízhaehti adi t’áh amíi mehéh dzaah nıgot’ǫ́ t’áh ats’eleh. Amíi t’ahsíi tsı̨ gųihthe meghǫnázhaeti k’eodı̨ ɂá saamba ts’ęhke mesaamba hule énidé dúle edi káondıh́ ǫt’e gots’ęh dúle thahne t’áh meghǫnázhaetı t’áh t’áh gots’áhodi: ats’eleh íle énidé t’ahsíi tśı̨ ts’ıhthé t’áh goghǫnázhaetı • Dzaah níots’éniɂǫ t’áh t’ahsí gots’ęh ts’enıdhę t’ah gǫndi k’ę́ ę́ ats’et’ı̨̨ gha góɂǫ íle énidé goghǫnázhaeti íle énidé godı̨ náts’edéh gokųę theɂǫ goch’á t’áh gogha gǫndi ts’ehtsi. -

Regulation GHA-RA, Maintenance of the Position Classification Plan
GHA-RA MONTGOMERY COUNTY REGULATION PUBLIC SCHOOLS Related Entries: Responsible Office: Human Resources and Development Maintenance of the Position Classification Plan I. PURPOSE To establish procedures for determining a position classification, reclassifying an existing position, reconstituting a position, classifying a proposed additional position, and writing and revising position specifications II. DEFINITIONS A. An appropriate officer is the superintendent of schools, deputy superintendent, chief operating officer, associate superintendent, department director, principal, or immediate supervisor. B. Classification/reclassification is a process to determine salary grades. A reclassification study involves identifying a significant change in the duties and responsibilities of a position that may require additional knowledge, skills, and abilities as a result of reorganization, new technologies, program revisions, and other events that impact the nature of the work to be performed that may or may not result in a salary grade change. C. Reconstitution involves assigning an existing or vacant position to a more appropriate classification and salary grade or the establishment of a new classification, typically as the result of reorganization or restructuring of an office or work unit. III. PROCEDURES A. Determination of a Position Classification Positions are included in the same classification when the same descriptive title applies, the same salary grade is equitable for all of the positions in the classification, and the following are substantially similar: 1 of 5 GHA-RA 1. Duties and responsibilities of the positions 2. Education and experience required 3. Tests or other methods used to determine applicants’ knowledge, abilities, and skills B. Reclassification of an Existing Position 1. Each December, the Office of Human Resources and Development (OHRD) with the approval of the chief operating officer sends a memorandum to the appropriate officers which outlines that year’s reclassification study process. -

GHA Fact Sheet Series Section 3 Program
For information: Justin Herter Public Information Officer (409) 765-1923 [email protected] GHA Fact Sheet Series Section 3 Program The GHA is currently assessing how to develop a Section 3 business and/or train and utilize Section 3 residents in its redevelopment initiatives. Since the storm, the GHA has developed a revitalization plan that includes the redevelopment of four public housing sites and up to 179 scattered site housing units throughout the island. These initiatives will be a combination of both new construction and rehabilitation of vacant structures in need of repair. By working to rebuild properties, the GHA will be in an incredible position to contribute to the economic recovery on the island as well as increase the affordable housing opportunities. Section Three of the Housing and Urban Development Act of 1968, as amended by the Housing & Community Development Act of 1994, requires that, to the greatest extent feasible, employment and other economic opportunities generated by HUD funds be directed to low and very low income residents. 24 CFR Part 135 requires that the GHA make best efforts to ensure that 30% of new hires of the GHA and of its contractors funded through development assistance, operating assistance or modernization assistance from HUD be residents of GHA communities. Furthermore, it requires that best efforts be made to ensure that 10% of all construction or repair related contracts and 3% of all other contracts be awarded to Section 3 business concerns. A Section Three Business Concern is defined as a business concern that is 51% owned by a GHA or other Section 3 residents, or 30% of whose permanent full time employees are GHA or other Section 3 residents or have been within the past three years. -

Symphony Orchestra
School of Music ROMANTIC SMORGASBORD Symphony Orchestra Huw Edwards, conductor Maria Sampen, violin soloist, faculty FRIDAY, OCT. 11, 2013 SCHNEEBECK CONCERT HALL 7:30 P.M. First Essay for Orchestra, Opus 12 ............................ Samuel Barber (1910–1983) Violin Concerto in D Major, Opus 77 .........................Johannes Brahms Allegro non troppo--cadenza--tranquillo (1833–1897) Maria Sampen, violin INTERMISSION A Shropshire Lad, Rhapsody for Orchestra ..................George Butterworth (1885–1916) Symphony No. 2 in D Major, Opus 43 ............................Jean Sibelius Allegro moderato--Moderato assai--Molto largamente (1865–1957) SYMPHONY ORCHESTRA Huw Edwards, conductor VIOLIN I CELLO FRENCH HORN Zachary Hamilton ‘15, Faithlina Chan ’16, Matt Wasson ‘14 concertmaster principal Billy Murphy ‘16 Marissa Kwong ‘15 Bronwyn Hagerty ‘15 Chloe Thornton ‘14 Jonathan Mei ‘16 Will Spengler ‘17 Andy Rodgers ‘16- Emily Brothers ‘14 Kira Weiss ‘17 Larissa Freier ‘17 Anna Schierbeek ‘16 TRUMPET Sophia El-Wakil ‘16 Aiden Meacham ‘14 Gavin Tranter ‘16 Matt Lam ‘16 Alana Roth ‘14 Lucy Banta ‘17 Linnaea Arnett ‘17 Georgia Martin ‘15 Andy Van Heuit ‘17 Abby Scurfield ‘16 Carolynn Hammen ‘16 TROMBONE VIOLIN II BASS Daniel Thorson ‘15 Clara Fuhrman ‘16, Kelton Mock ‘15 Stephen Abeshima ‘16 principal principal Wesley Stedman ‘16 Rachel Lee ‘15 Stephen Schermer, faculty Sophie Diepenheim ‘14 TUBA Brandi Main ‘16 FLUTE and PICCOLO Scott Clabaugh ‘16 Nicolette Andres ‘15 Whitney Reveyrand ‘15 Lauren Griffin ‘17 Morgan Hellyer ‘14 TIMPANI and -

Keith Phares, Baritone, and Michael Baitzer, Piano Department of Music, University of Richmond
University of Richmond UR Scholarship Repository Music Department Concert Programs Music 3-21-2005 Keith Phares, baritone, and Michael Baitzer, piano Department of Music, University of Richmond Follow this and additional works at: https://scholarship.richmond.edu/all-music-programs Part of the Music Performance Commons Recommended Citation Department of Music, University of Richmond, "Keith Phares, baritone, and Michael Baitzer, piano" (2005). Music Department Concert Programs. 342. https://scholarship.richmond.edu/all-music-programs/342 This Program is brought to you for free and open access by the Music at UR Scholarship Repository. It has been accepted for inclusion in Music Department Concert Programs by an authorized administrator of UR Scholarship Repository. For more information, please contact [email protected]. Monday, March 21, 2005 • 7:30pm Modlin Center for the Arts Call),p Concert Hall, Booker Hall of Music Keith Phares, baritone Michael Baitzer, piano UN!VER.SlTY OF RICHMOND Sponsored in part by the 175th Anniversary Committee and the University's Cultural Affairs Committee The Modlin Center thanks Style Weekly for media sponsorship of the 2004-2005 season. Tonight's Program La bonne chanson, Op. 61 (Paul Verlaine) Gabriel Faure "Une Sainte en son aureole" (1845-1924) "Puisque l'aube gpndit" "La June blanche luit dans les bois" "J'allais par des chemins perfides" "J'ai presque peur, en verite" "Avant que tune t'en ailles" "Done, ce sera par un clair jour d'ete" "N'est-ce pas?" "L:hiver a cesse" Lieder eines fahrenden Gesellen Gustav Mahler "Wenn mein Schatz Hochzeit macht" (1860-1911) "Gieng heut' Morgen tiber's Feld" "Ich hab' ein gltiend Messer" "Die zwei blauen Augen" -Intermission- Bredon Hill and Other Songs (A.E. -

Georgia Coverdell Acute Stroke Registry Quarterly Newsletter WINTER 2013 Coverdell Partners: Georgia Coverdell 2013 Awards
Georgia Coverdell Acute Stroke Registry Quarterly Newsletter WINTER 2013 Coverdell Partners: Georgia Coverdell 2013 Awards Georgia Department of Public Health “Georgia Coverdell Champion Hospital of the Year” (DPH) Award Winners Emory University School of Medicine CONGRATULATIONS TO THE FOLLOWING 4 HOSPITALS: Georgia Medical Care Foundation Putnam General Hospital (very small hospital, 25 beds or less) (GMCF) American Stroke Dekalb Medical Hillandale (small hospital, 26–100 beds) Association (ASA) Georgia Hospital South Georgia Medical Center (medium hospital, 101–350 beds) Association (GHA) Grady Health System (large hospital, over 350 beds) If you have anything you would All Georgia Coverdell hospitals have the opportunity to receive the award, which is like included in an based on a point system. A total of four hospitals receiving the most points during upcoming the period from January 1, 2013 through December 31, 2013 will be the recipients newsletter or have of next year’s award. The award is a framed certificate to share with your hospital achieved recent and hospital administration. recognition in the area of stroke, The point system is based on the following criteria: contact: Participation Points Allotted Hospital attendance on GA Monthly Coverdell Call 1 Kerrie Krompf Hospital presenting on GA Monthly Coverdell Call 5 [email protected] Physician Champion presenting on GA Monthly Coverdell Call 10 Published Q-Tip in Coverdell Quarterly Newsletter 5 or Published Article in Coverdell Quarterly Newsletter 10 Published “Blurb” (250 words) in Coverdell Quarterly Newsletter 2 770-380-8998 Published Stroke Survivor Story in Coverdell Quarterly Newsletter 10 Workshop attendance (per hospital) 5 In the event that multiple articles, blurbs, stroke survivor stories, Q-tips, etc. -

GHA Letterhead Template
Depression Screening Background As many as 1 in 6 people over 65 suffer from depression. Estimates show depression occurs in 25% of older Americans with other illness, such as: • Cancer • Arthritis • Stroke • Chronic lung disease • Cardiovascular disease • Chronic pain Factors such as loss of loved ones or friends increase the likelihood of depression. Health care providers miss opportunities to improve mental health and general medical outcomes when mental illness is under-recognized and undertreated in primary care settings. Depression Screening Utilization Data analysis shows WPS GHA providers in Jurisdiction 5 (J5) and Jurisdiction 8 (J8) are underutilizing depression screening. The 2019 rate of depression screening ranges from 1.8% to 6.4% of beneficiaries. See below for specific state data. The average screening rate of all beneficiaries is: • Jurisdiction 5 (J5) is about 2.8% • Jurisdiction 8 (J8) is about 3.8% • National screening rate is 5.44% for all Medicare Administrative Contractors (MACs). Providers in J5 and J8 underutilize depression screening, while the prevalence of depression is relatively high. The range is from 17% in Nebraska to 20.4% in Indiana and Missouri. The rates of all states except for that of Nebraska, are higher than the national rate of 17.9%. Depression Screening Coverage Criteria The following coverage criteria apply: • Anyone eligible for Medicare • Screening only. It does not apply to: o Depression treatment o Chronic condition caused by depression • One year between screenings o 11 full months must pass following the month in which the screening occurred o Example: depression screening on 7/12/2019, next available 7/1/2020 • Furnished in a primary care setting with staff assisted depression care to ensure o Accurate diagnosis o Effective treatment o Follow-up • Up to 15-minute screening o This includes time spent to: . -
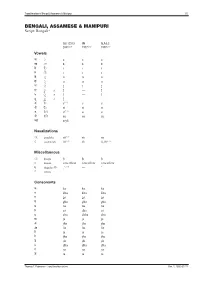
Transliteration of <Script Name>
Transliteration of Bengali, Assamese & Manipuri 1/5 BENGALI, ASSAMESE & MANIPURI Script: Bengali* ISO 15919 UN ALA-LC 2001(1.0) 1977(2.0) 1997(3.0) Vowels অ ◌ a a a আ ◌া ā ā ā ই ি◌ i i i ঈ ◌ী ī ī ī উ ◌ু u u u ঊ ◌ূ ū ū ū ঋ ◌ৃ r̥ ṛ r̥ ৠ ◌ৄ r̥̄ — r̥̄ ঌ ◌ৢ l̥ — l̥ ৡ ◌ৣ A l̥ ̄ — — এ ে◌ e(1.1) e e ঐ ৈ◌ ai ai ai ও ে◌া o(1.1) o o ঔ ে◌ৗ au au au অ�া a:yā — — Nasalizations ◌ং anunāsika ṁ(1.2) ṁ ṃ ◌ঁ candrabindu m̐ (1.2) m̐ n̐, m̐ (3.1) Miscellaneous ◌ঃ bisarga ḥ ḥ ḥ ◌্ hasanta vowelless vowelless vowelless (1.3) ঽ abagraha Ⓑ :’ — ’ ৺ isshara — — — Consonants ক ka ka ka খ kha kha kha গ ga ga ga ঘ gha gha gha ঙ ṅa ṅa ṅa চ ca cha ca ছ cha chha cha জ ja ja ja ঝ jha jha jha ঞ ña ña ña ট ṭa ṭa ṭa ঠ ṭha ṭha ṭha ড ḍa ḍa ḍa ঢ ḍha ḍha ḍha ণ ṇa ṇa ṇa ত ta ta ta Thomas T. Pedersen – transliteration.eki.ee Rev. 2, 2005-07-21 Transliteration of Bengali, Assamese & Manipuri 2/5 ISO 15919 UN ALA-LC 2001(1.0) 1977(2.0) 1997(3.0) থ tha tha tha দ da da da ধ dha dha dha ন na na na প pa pa pa ফ pha pha pha ব ba ba ba(3.2) ভ bha bha bha ম ma ma ma য ya ja̱ ya র ⒷⓂ ra ra ra ৰ Ⓐ ra ra ra ল la la la ৱ ⒶⓂ va va wa শ śa sha śa ষ ṣa ṣha sha স sa sa sa হ ha ha ha ড় ṛa ṙa ṛa ঢ় ṛha ṙha ṛha য় ẏa ya ẏa জ় za — — ব় wa — — ক় Ⓑ qa — — খ় Ⓑ k̲h̲a — — গ় Ⓑ ġa — — ফ় Ⓑ fa — — Adscript consonants ◌� ya-phala -ya(1.4) -ya ẏa ৎ khanda-ta -t -t ṯa ◌� repha r- r- r- ◌� baphala -b -b -b ◌� raphala -r -r -r Vowel ligatures (conjuncts)C � gu � ru � rū � Ⓐ ru � rū � śu � hr̥ � hu � tru � trū � ntu � lgu Thomas T. -

Balinese Romanization Table
Balinese Principal consonants1 (h)a2 ᬳ ᭄ᬳ na ᬦ ᭄ᬦ ca ᬘ ᭄ᬘ᭄ᬘ ra ᬭ ᭄ᬭ ka ᬓ ᭄ᬓ da ᬤ ᭄ᬤ ta ᬢ ᭄ᬢ sa ᬲ ᭄ᬲ wa ᬯ la ᬮ ᭄ᬙ pa ᬧ ᭄ᬧ ḍa ᬟ ᭄ᬠ dha ᬥ ᭄ᬟ ja ᬚ ᭄ᬚ ya ᬬ ᭄ᬬ ña ᬜ ᭄ᬜ ma ᬫ ᭄ᬫ ga ᬕ ᭄ᬕ ba ᬩ ᭄ᬩ ṭa ᬝ ᭄ᬝ nga ᬗ ᭄ᬗ Other consonant forms3 na (ṇa) ᬡ ᭄ᬡ ca (cha) ᭄ᬙ᭄ᬙ ta (tha) ᬣ ᭄ᬣ sa (śa) ᬱ ᭄ᬱ sa (ṣa) ᬰ ᭄ᬰ pa (pha) ᬨ ᭄ᬛ ga (gha) ᬖ ᭄ᬖ ba (bha) ᬪ ᭄ᬪ ‘a ᬗ᬴ ha ᬳ᬴ kha ᬓ᬴ fa ᬧ᬴ za ᬚ᬴ gha ᬕ᬴ Vowels and other agglutinating signs4 5 a ᬅ 6 ā ᬵ ᬆ e ᬾ ᬏ ai ᬿ ᬐ ĕ ᭂ ö ᭃ i ᬶ ᬇ ī ᬷ ᬈ o ᭀ ᬑ au ᭁ ᬒ u7 ᬸ ᬉ ū ᬹ ᬊ ya, ia8 ᭄ᬬ r9 ᬃ ra ᭄ᬭ rĕ ᬋ rö ᬌ ᬻ lĕ ᬍ ᬼ lö ᬎ ᬽ h ᬄ ng ᬂ ng ᬁ Numerals 1 2 3 4 5 ᭑ ᭒ ᭓ ᭔ ᭕ 6 7 8 9 0 ᭖ ᭗ ᭘ ᭙ ᭐ 1 Each consonant has two forms, the regular and the appended, shown on the left and right respectively in the romanization table. The vowel a is implicit after all consonants and consonant clusters and should be supplied in transliteration, unless: (a) another vowel is indicated by the appropriate sign; or (b) the absence of any vowel is indicated by the use of an adeg-adeg sign ( ). (Also known as the tengenen sign; ᭄ paten in Javanese.) 2 This character often serves as a neutral seat for a vowel, in which case the h is not transcribed. -

Nonatonic Harmonic Structures in Symphonies by Ralph Vaughan Williams and Arnold Bax Cameron Logan [email protected]
University of Connecticut OpenCommons@UConn Doctoral Dissertations University of Connecticut Graduate School 12-2-2014 Nonatonic Harmonic Structures in Symphonies by Ralph Vaughan Williams and Arnold Bax Cameron Logan [email protected] Follow this and additional works at: https://opencommons.uconn.edu/dissertations Recommended Citation Logan, Cameron, "Nonatonic Harmonic Structures in Symphonies by Ralph Vaughan Williams and Arnold Bax" (2014). Doctoral Dissertations. 603. https://opencommons.uconn.edu/dissertations/603 i Nonatonic Harmonic Structures in Symphonies by Ralph Vaughan Williams and Arnold Bax Cameron Logan, Ph.D. University of Connecticut, 2014 This study explores the pitch structures of passages within certain works by Ralph Vaughan Williams and Arnold Bax. A methodology that employs the nonatonic collection (set class 9-12) facilitates new insights into the harmonic language of symphonies by these two composers. The nonatonic collection has received only limited attention in studies of neo-Riemannian operations and transformational theory. This study seeks to go further in exploring the nonatonic‟s potential in forming transformational networks, especially those involving familiar types of seventh chords. An analysis of the entirety of Vaughan Williams‟s Fourth Symphony serves as the exemplar for these theories, and reveals that the nonatonic collection acts as a connecting thread between seemingly disparate pitch elements throughout the work. Nonatonicism is also revealed to be a significant structuring element in passages from Vaughan Williams‟s Sixth Symphony and his Sinfonia Antartica. A review of the historical context of the symphony in Great Britain shows that the need to craft a work of intellectual depth, simultaneously original and traditional, weighed heavily on the minds of British symphonists in the early twentieth century.