A Novel Approach to Dream Content Analysis Reveals Links Between Learning-Related Dream Incorporation and Cognitive Abilities
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Nature of Delayed Dream Incorporation ('Dream-Lag Effect')
Received: 19 December 2017 | Revised: 20 March 2018 | Accepted: 21 March 2018 DOI: 10.1111/jsr.12697 REGULAR RESEARCH PAPER The nature of delayed dream incorporation (‘dream-lag effect’): Personally significant events persist, but not major daily activities or concerns Jean-Baptiste Eichenlaub1 | Elaine van Rijn1 | Mairead Phelan1 | Larnia Ryder1 | M. Gareth Gaskell2 | Penelope A. Lewis3 | Matthew P. Walker4 | Mark Blagrove1 1Swansea University Sleep Laboratory, Department of Psychology, Swansea Summary University, Swansea, UK Incorporation of details from waking life events into rapid eye movement (REM) 2Sleep, Language and Memory Laboratory, sleep dreams has been found to be highest on the 2 nights after, and then 5–7 Department of Psychology, University of York, York, UK nights after, the event. These are termed, respectively, the day-residue and dream- 3School of Psychology and Cardiff lag effects. This study is the first to categorize types of waking life experiences and University Brain Imaging Centre, Cardiff University, Cardiff, UK compare their incorporation into dreams across multiple successive nights. Thirty- 4Center for Human Sleep Science, eight participants completed a daily diary each evening and a dream diary each Department of Psychology, University of morning for 14 days. In the daily diary, three categories of experiences were California, Berkeley, CA, USA reported: major daily activities (MDAs), personally significant events (PSEs) and Correspondence major concerns (MCs). After the 14-day period each participant identified -

What Is Sleep?
What Is Sleep? By SIRI HUSTVEDT The New York Times, April 21, 2010, 9:00 pm I am convinced that during bouts of insomnia I have sometimes slept without knowing it. The thoughts of waking seem to mingle with thoughts that may be part of sleep. Has the clock moved too quickly? Did I doze off? Some years ago in a rented house in Vermont, I couldn’t sleep and lay awake listening to the sounds of mice in the walls, bears that sounded like owls calling to each other in the woods and the wind in the trees. I then dreamed I was lying awake on the very bed where in fact I was sleeping, but someone had broken into the house. Because the room where I actually was and the room I dreamed were identical, the threshold between waking and sleeping had blurred and, when I woke up, I thought I heard the burglar moving around downstairs. It was a frightening experience, a temporary loss of the boundaries between waking experience and the illusions of dreams. Once, my sister Ingrid, while lying on her bed, certain she was fully awake, was amazed to see a strange man wheel a bicycle into her bedroom. After a minute or so, the man and his vehicle disappeared. They had been either a dream or a hallucination. As with me, my sister’s confusion of the real and the unreal turned on the fact that both were located in the same place. In his “Meditations,” Rene Descartes asked if he could be really certain he was awake. -

Lucid Dreaming and Personality in Children/Ado- Lescents and Adults
Brief report I J o D R Lucid dreaming and personality in children/ado- lescents and adults: The UK library study Michael Schredl1, Josie Henley-Einion2, & Mark Blagrove2 1Central Institute of Mental Health, Medical Faculty Mannheim, Heidelberg University, Ger- many 2Department of Psychology, Swansea University, United Kingdom Summary. Research that has focused on the relationship between the Big Five personality dimensions and lucid dream- ing frequency has been restricted to student samples. The present study included adolescents and adults (N = 1375). i.e., the sample included a large range of ages. Lucid dreaming was more strongly related to openness to experiences com- pared to previous findings. The small but significant negative correlation between conscientiousness and lucid dreaming should be followed up by studies relating the Big Five personality factors to the contents of lucid dreams. Keywords: Lucid dreaming, personality 1. Introduction whereas Schredl and Erlacher (2004) found that not the total openness score showed a significant relationship to lucid The term lucid dream is defined as a dream in which the dreaming but solely the two openness to experience facets dreamer – while dreaming – is aware that she/he is dream- (“fantasy”, “ideas”). It has to be mentioned that these stud- ing (LaBerge & Rheingold, 1990; Tholey & Utecht, 1987). ies (Schredl & Erlacher, 2004; Watson, 2001) were carried Within the lucid dream the dreamer can control some of out in student samples. the events or content of the dream (LaBerge, 1985). Lucid The aim of the following study is to examine the relation- dreaming can be a useful application for the training of skills ship between the Big Five personality dimensions and lucid (Erlacher & Schredl, 2010; Stumbrys, Erlacher, & Schredl, dreaming frequency in a sample with a large age range that 2016) and help to cope with nightmares (Brylowski, 1990; includes adolescents and adults. -

Conference Organized by Itzhak Fried (UCLA / Tel-Aviv University) As Part of the Paris IAS Brain, Culture & Society Program
Conference organized by Itzhak Fried (UCLA / Tel-Aviv University) as part of the Paris IAS Brain, Culture & Society Program From an interdisciplinary perspective including neuroscience, medicine, the humanities and art, the meeting aims at (1) advancing and disseminating scientific knowledge on how specific sleep processes aid memory consolidation (2) inspiring science and arts to adopt new approaches to the importance of sleep and dreams (3) benefiting society by promoting awareness for good sleep habits and their effect on cognitive well-being. Cognition during Sleep Unsuspected cognition in the sleeping brain Sid Kouider (CNRS - ENS) Sleep has been argued to be the price to pay for neural plasticity: it allows optimising memory consolidation at the price of rendering organisms vulnerable to external threats. Yet, recent research reveals that the sleeping brain is actually not fully shut down from the environment, as it continues registering and integrating external events to some extent. This raises the questions of why would sleepers continue processing external information and why do they remain unresponsive at the behavioural level? Here I will argue that the sleeping brain attempts to finely balance the need to turn inward in order to optimise memory consolidation with the ability to rapidly revert to wakefulness when necessary. This leads to the hypothesis that sleepers enter a “standby mode” in which neural mechanisms aimed at tracking relevant signals in the environment remain functional. I will present several studies using neural markers of cognitive processing to show that the human brain, even after falling asleep, continues to 1) classify auditory events in a task-dependent manner, 2) rely on selective attention to resolve the cocktail party phenomenon, and 3) even form new memory contents on perpetual learning tasks. -

Sleep and Dreaming: Scientific Advances and Reconsiderations Edited by Edward F
Cambridge University Press 0521810442 - Sleep and Dreaming: Scientific Advances and Reconsiderations Edited by Edward F. Pace-Schott, Mark Solms, Mark Blagrove and Stevan Harnad Frontmatter More information Sleep and Dreaming How and why does the sleeping brain generate dreams? Though the question is old, a paradigm shift is now oc- curring in the science of sleep and dreaming that is making room for new answers. From brainstem-based models of sleep cycle control, research is moving toward combined brainstem/forebrain models of sleep cognition itself. Fur- thermore, advances in philosophy, psychiatry, psychology, artificial intelligence, neural network modeling, psycho- physiology, neurobiology, and clinical medicine make this a propitious time to review and bridge the gaps among these fields as they relate to sleep and dream research. This book presents five papers by leading scientists at the center of the current firmament and more than seventy-five commentaries on those papers by nearly all the other lead- ing authorities in the field. Topics include mechanisms of dreaming and REM sleep, memory consolidation in REM sleep, and an evolutionary hypothesis of the function of dreaming. The papers and commentaries, together with the authors’ rejoinders, represent a huge leap forward in our understanding of the sleeping and dreaming brain, ulti- mately offering new and unique views of consciousness and cognition. They help provide new answers to both old and new questions, based on the latest findings in modern brain research. The book’s multidisciplinary perspective will ap- peal to students and researchers in neuroscience, cognitive science, and psychology. Edward F. Pace-Schott is Instructor in Psychiatry at Har- vard Medical School. -

FALL 2019 in Progress.Indd
Mark Blagrove is Professor of Psychology at Swansea University, Wales, and Director of the Sleep Laboratory at Swansea University. Dr Julia Lockheart is Senior Lecturer at Swansea College of Art, University of Wales Trinity St David, and Associate Lecturer at Goldsmiths, University of London. Their collaboration can be seen on the website DreamsID.com. uring the wonderful Rolduc conference I in the audience. The background to the second (Mark Blagrove) was asked to write an article incident was that the scientists generally didn’t Ddescribing how a sleep scientist got involved go to experiential sessions back then. So it was in an arts science collaboration, with artist Julia eye-opening when Milton Kramer emerged Lockheart, and the role of IASD in that. It’s a long from a Gayle Delaney workshop session story! and told many of us that Gayle’s exploration of a single dream was done so well, and so I fi rst joined what was the Association for the Study of carefully, leading to so much information and Dreams in 1989, and went to my fi rst ASD conference insight about and for the dream sharer. that year, in London. I had just fi nished my PhD, on the relationship of dream content to waking life Eventually, we were to have scientist Michael cognition. Before that I had studied experimental Schredl running one of the morning dream psychology as part of my Natural Sciences degree at groups. I also came to do so, but by an Cambridge, and had got very interested in dreaming. unexpected route. I had started to attend The London conference was wonderful, meeting so morning dream groups at the conference, many famous scientists, including Ros Cartwright, partly to see what could be done with such Ernest Hartmann, Milton Kramer and Allan Hobson. -

Contents More Information
Cambridge University Press 0521810442 - Sleep and Dreaming: Scientific Advances and Reconsiderations Edited by Edward F. Pace-Schott, Mark Solms, Mark Blagrove and Stevan Harnad Table of Contents More information Contents Preface page ix M. Blagrove Introduction xi 1 J. A. Hobson, E. F. Pace-Schott, and R. Stickgold Dreaming and the brain: Toward a cognitive neuroscience of conscious states 1 2 M. Solms Dreaming and REM sleep are controlled by different brain mechanisms 51 3 T. A. Nielsen A review of mentation in REM and NREM sleep: “Covert” REM sleep as a possible reconciliation of two opposing models 59 4 R. P. Vertes and K. E. Eastman The case against memory consolidation in REM sleep 75 5 A. Revonsuo The reinterpretation of dreams: An evolutionary hypothesis of the function of dreaming 85 Open Peer Commentary and Authors’ Responses Table of Commentators 112 Open Peer Commentary Antrobus, J. S. How does the dreaming brain explain the dreaming mind? 115 Ardito, R. B. Dreaming as an active construction of meaning 118 Bednar, J. A. Internally-generated activity, non-episodic memory, and emotional salience in sleep 119 Blagrove, M. Dreams have meaning but no function 121 Borbély, A. A. & Wittmann, L. Sleep, not REM sleep, is the royal road to dreams 122 Born, J. & Gais, S. REM sleep deprivation: The wrong paradigm leading to wrong conclusions 123 Bosinelli, M. & Cicogna, P. C. REM and NREM mentation: Nielsen’s model once again supports the supremacy of REM 124 Cartwright, R. How and why the brain makes dreams: A report card on current research on dreaming 125 Cavallero, C. -

Characteristics of the Memory Sources of Dreams: a New Version of the Content- Matching Paradigm to Take Mundane and Remote Memories Into Account
RESEARCH ARTICLE Characteristics of the memory sources of dreams: A new version of the content- matching paradigm to take mundane and remote memories into account Raphael Vallat1, Benoit Chatard1, Mark Blagrove2, Perrine Ruby1* 1 Lyon Neuroscience Research Center (CRNL), Brain Dynamics and Cognition Team (DYCOG), INSERM UMRS 1028, CNRS UMR 5292, Universite Claude Bernard Lyon 1, Universite de Lyon, Lyon, France, 2 Swansea University, Sleep laboratory, Swansea, Wales, United Kingdom a1111111111 a1111111111 * [email protected] a1111111111 a1111111111 a1111111111 Abstract Several studies have demonstrated that dream content is related to the waking life of the dreamer. However, the characteristics of the memory sources incorporated into dreams are OPEN ACCESS still unclear. We designed a new protocol to investigate remote memories and memories of Citation: Vallat R, Chatard B, Blagrove M, Ruby P trivial experiences, both relatively unexplored in dream content until now. Upon awakening, (2017) Characteristics of the memory sources of for 7 days, participants identified the waking life elements (WLEs) related to their dream con- dreams: A new version of the content-matching tent and characterized them and their dream content on several scales to assess notably paradigm to take mundane and remote memories emotional valence. Thanks to this procedure, they could report WLEs from the whole life- into account. PLoS ONE 12(10): e0185262. https:// doi.org/10.1371/journal.pone.0185262 span, and mundane ones before they had been forgotten. Participants (N = 40, 14 males, age = 25.2 7.6) reported 6.2 2.0 dreams on average. For each participant, 83.4% 17.8 Editor: Raffaele Ferri, Associazione OASI Maria SS, ± ± ± ITALY of the dream reports were related to one or more WLEs. -

Nightmares, Clues to Why We Dream at All
October 23, 2007 BASICS In the Dreamscape of Nightmares, Clues to Why We Dream at All By NATALIE ANGIER The patient was a 37-year-old man who had been physically abused as a boy by his schizophrenic mother, often while he lay in bed trying to fall asleep. Nevertheless, he had grown into a reasonably normal, gainfully employed adult, and he thought that the worst was behind him, until one night he awoke to find an intruder rummaging through his dresser drawers. After that, his nightmares began — terrifying, recurrent dreams in which the intruder was a middle-age woman and a knife dangled with Damoclesian contempt from the ceiling fan over his head. “The old fear memories had not gone away,” said Dr. Ross Levin, a psychologist and sleep researcher at Yeshiva University in New York. They “were easily reactivated by the recent trauma,” and just as readily twisted into the basis of a repetitive nightmare. Dr. Levin urged the patient to reframe the dream and rehearse alternatives to swinging blades and frozen fear, until finally the nightmares abated and the man could regain his footing. Few of us suffer from nightmares crippling and persistent enough to demand treatment. Yet we all know how bad a nightmare feels, how it surrounds you and surges up to drown you and makes your teeth fall out in chunks and gives you leukemia and look, your 6-year-old daughter is running back and forth through traffic, and oh no, this train is headed the wrong way and it’s past midnight, and there you are a cowardly third-grader back on Creston Avenue in the Bronx, no, please, not the Bronx! And you scream and you thrash and you want to wake up. -
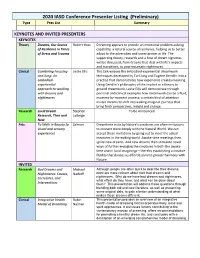
2020 IASD Conference Presenter Listing (Preliminary) Type Pres List Summary
2020 IASD Conference Presenter Listing (Preliminary) Type Pres List Summary KEYNOTES AND INVITED PRESENTERS KEYNOTES Theory Dreams, Our Source Robert Hoss Dreaming appears to provide an emotional problem-solving of Resilience in Times capability, a natural source of resilience, helping us to better of Stress and Trauma adapt to the adversities and uncertainties in life. The supporting theory, research and a host of dream vignettes will be discussed, from dreams that deal with life’s impacts and transitions, to post-traumatic nightmares. Clinical Combining Focusing Leslie Ellis This talk weaves the embodied-experiential dreamwork and Jung: An techniques developed by Carl Jung and Eugene Gendlin into a embodied- practice that demonstrates how experience creates meaning. experiential Using Gendlin’s philosophy of the implicit as a theory to approach to working ground dreamwork, Leslie Ellis will demonstrate through with dreams and personal and clinical examples how dreamwork can be a fluid, nightmares moment-by-moment process; a certain kind of attention invites dreams to shift into waking imaginal journeys that bring fresh perspectives, insight and change. Research Lucid Dream Stephen To Be Announced Research, Then and LaBerge Now Arts To Walk in Beauty (a Salmon Dreamtime visits by Nature’s creatures are often invitations visual and sensory to connect more deeply with the Natural World. We can experience) accept these invitations by going out to meet the actual creatures in the waking world. Awake-time meetings then ignite new dreams. And new dreams then stimulate novel ways of further engaging the creatures in both the awake- time and in lucid imaginings—thereby establishing a creative fluidity that dances us effortlessly into greater harmony with Nature. -

Professor Mark Blagrove Swansea University Sleep Laboratory Department of Psychology Swansea University SA2 0LY UK
Os textos são da exclusiva responsabilidade dos autores All texts are of the exclusive responsibility of the authors Electrophysiological correlates of the incorporation of recent memory sources into REM and non-REM dreams and of levels of insight following REM and non-REM dream interpretation ABSTRACT: Background There have been claims throughout history that the consideration of dreams can result in personal realizations and insight. Aim To assess insight resulting from discussing Rapid Eye Movement (REM) and non-REM stage 2 (N2) dreams collected in the sleep laboratory, with discussion of daydreams as a control, and electrophysiological correlates of insight. Method 31 participants slept in the sleep laboratory and were awakened for REM and N2 dream reports, and cued for a daydream report. Participants discussed their dream and daydream reports with experimenters who had not been involved in collecting the dreams and daydreams, and scored the discussions for insight obtained. Results There were no insight differences between discussing REM and N2 dreams. Discussing dreams led to higher scores on the exploration-insight measure than did discussing daydreams. The differences were related to the learning of what the report means in terms of waking life issues. Conclusions Despite electrophysiological and functional differences between REM and N2 sleep, dreams from these stages did not differ in exploration-insight scores, but did exceed exploration-insight for the discussion of daydreams. Daydreams did not show the memory-consolidation related delayed incorporation of waking life events that has been found for REM dreams (van Rijn et al., 2016, 2018). A second main paper is under review, with preliminary results in Blagrove et al. -

Lucid Dreaming in Children: the UK Library Study
Lucid dreaming in children I J o D R Lucid dreaming in children: The UK library study Michael Schredl1, Josie Henley-Einion2 and Mark Blagrove2 1Central Institute of Mental Health, Mannheim, Germany 2Department of Psychology, Swansea University, United Kingdom Summary. The term lucid dream designates a dream in which the dreamer, while dreaming, is aware he or she is dream- ing. Whereas lucid dreaming has been studied in adults, large-scaled surveys in children are scarce. The sample here included 3579 children for the ages from 6 to 18 who completed a brief questionnaire distributed in UK libraries. 43.5% reported having had at least one lucid dream. As hypothesized, incidence of lucid dreams was most strongly associated with the preference for reading fantasy/science-fiction. Whether lucid dreaming might be beneficial for children in skills training or coping with nightmares are interesting topics for future research. Keywords: Lucid dreaming, Children, Book preferences 1. Introduction experiences and after several steps in cognitive develop- ment (around the age of 9 or 10), the child understands that The term lucid dream designates a dream in which the dreams are internally produced images during sleep. More dreamer, while dreaming, is aware he or she is dreaming recent studies (Wilson, 1991; Meyer & Shore, 2001), howev- (Tholey, 1983; LaBerge & Rheingold, 1990). In student sam- er, showed that most five-year olds do perceive dreams as ples, the percentage of people who report having had at internal mental processes. Yet the concept of lucid dreaming least one lucid dream in their lifetime ranges from 57.5% might be more difficult to understand: a study in preschool (Gackenbach, 1991) to 73% (Blackmore, 1982) and 82% children (4 to 6 years) found that most children believe that (Schredl & Erlacher, 2004).