Plant Biodiversity
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Heterospory: the Most Iterative Key Innovation in the Evolutionary History of the Plant Kingdom
Biol. Rej\ (1994). 69, l>p. 345-417 345 Printeii in GrenI Britain HETEROSPORY: THE MOST ITERATIVE KEY INNOVATION IN THE EVOLUTIONARY HISTORY OF THE PLANT KINGDOM BY RICHARD M. BATEMAN' AND WILLIAM A. DiMlCHELE' ' Departments of Earth and Plant Sciences, Oxford University, Parks Road, Oxford OXi 3P/?, U.K. {Present addresses: Royal Botanic Garden Edinburiih, Inverleith Rojv, Edinburgh, EIIT, SLR ; Department of Geology, Royal Museum of Scotland, Chambers Street, Edinburgh EHi ijfF) '" Department of Paleohiology, National Museum of Natural History, Smithsonian Institution, Washington, DC^zo^bo, U.S.A. CONTENTS I. Introduction: the nature of hf^terospon' ......... 345 U. Generalized life history of a homosporous polysporangiophyle: the basis for evolutionary excursions into hetcrospory ............ 348 III, Detection of hcterospory in fossils. .......... 352 (1) The need to extrapolate from sporophyte to gametophyte ..... 352 (2) Spatial criteria and the physiological control of heterospory ..... 351; IV. Iterative evolution of heterospory ........... ^dj V. Inter-cladc comparison of levels of heterospory 374 (1) Zosterophyllopsida 374 (2) Lycopsida 374 (3) Sphenopsida . 377 (4) PtiTopsida 378 (5) f^rogymnospermopsida ............ 380 (6) Gymnospermopsida (including Angiospermales) . 384 (7) Summary: patterns of character acquisition ....... 386 VI. Physiological control of hetcrosporic phenomena ........ 390 VII. How the sporophyte progressively gained control over the gametophyte: a 'just-so' story 391 (1) Introduction: evolutionary antagonism between sporophyte and gametophyte 391 (2) Homosporous systems ............ 394 (3) Heterosporous systems ............ 39(1 (4) Total sporophytic control: seed habit 401 VIII. Summary .... ... 404 IX. .•Acknowledgements 407 X. References 407 I. I.NIRODUCTION: THE NATURE OF HETEROSPORY 'Heterospory' sensu lato has long been one of the most popular re\ie\v topics in organismal botany. -

Comparative Transcriptome Analysis Suggests Convergent Evolution Of
Alejo-Jacuinde et al. BMC Plant Biology (2020) 20:468 https://doi.org/10.1186/s12870-020-02638-3 RESEARCH ARTICLE Open Access Comparative transcriptome analysis suggests convergent evolution of desiccation tolerance in Selaginella species Gerardo Alejo-Jacuinde1,2, Sandra Isabel González-Morales3, Araceli Oropeza-Aburto1, June Simpson2 and Luis Herrera-Estrella1,4* Abstract Background: Desiccation tolerant Selaginella species evolved to survive extreme environmental conditions. Studies to determine the mechanisms involved in the acquisition of desiccation tolerance (DT) have focused on only a few Selaginella species. Due to the large diversity in morphology and the wide range of responses to desiccation within the genus, the understanding of the molecular basis of DT in Selaginella species is still limited. Results: Here we present a reference transcriptome for the desiccation tolerant species S. sellowii and the desiccation sensitive species S. denticulata. The analysis also included transcriptome data for the well-studied S. lepidophylla (desiccation tolerant), in order to identify DT mechanisms that are independent of morphological adaptations. We used a comparative approach to discriminate between DT responses and the common water loss response in Selaginella species. Predicted proteomes show strong homology, but most of the desiccation responsive genes differ between species. Despite such differences, functional analysis revealed that tolerant species with different morphologies employ similar mechanisms to survive desiccation. Significant functions involved in DT and shared by both tolerant species included induction of antioxidant systems, amino acid and secondary metabolism, whereas species-specific responses included cell wall modification and carbohydrate metabolism. Conclusions: Reference transcriptomes generated in this work represent a valuable resource to study Selaginella biology and plant evolution in relation to DT. -

Desiccation Tolerance: Phylogeny and Phylogeography
Desiccation Tolerance: Phylogeny and Phylogeography BioQUEST Workshop 2009 Resurrection Plants Desiccation tolerant Survive dehydration Survive in dormant state for extended time period Survive rehydration Xerophyta humilis, from J. Farrant web site (http://www.mcb.uct.ac.za/Staff/JMF/index.htm) Figure 2. Selaginella lepidophylla http://en.wikipedia.org/wiki/ File:Rose_of_Jericho.gif Desiccation Issues Membrane integrity Protein structure Generation of free radicals Rascia, N, La Rocca, N. 2005 Critical Reviews in Plant Sciences Solutions Repair upon hydration Prevent damage during dehydration Production/accumulation of replacement solutes Inhibition of photosynthesis http://www.cbs.dtu.dk/staff/dave/roanoke/elodeacell.jpg Evolution of Desiccation Tolerance From Oliver, et al 2000. Plant Ecology Convergent Evolution Among Vascular Plants From Oliver, et al 2000. Plant Ecology Desiccation Tolerance in Vascular Plants Seeds Pollen Spores Osmotic stress Desertification 40% of Earth’s surface 38% of population Low productivity Lack of water Depletion of soil Loss of soil Delicate system Using Evolutionary Relationships to Identify Genes for Crop Enhancement If desiccation sensitive plants have retained genes involved in desiccation tolerance, perhaps expression of those genes can be genetically modified to enhance desiccation tolerance. Expression patterns in dehydratingTortula and Xerophyta Michael Luth Xerophyta humilis, from J. Farrant web site (http://www.mcb.uct.ac.za/Staff/JMF/index.htm) ? Methods Occurrence data for Anastatica hierochuntic: o Downloaded from the Global Biodiversity Information Facility (gbif.org). o Of 127 available occurrence points, only 68 were georeferenced with latitude and longitude information. Environmental layers: Precipitation and temperature from the IPCC climate dataset Niche Modeling: Maximum Entropy (Maxent) Maxent principle is to estimate the probability distribution such as the spatial distribution of a species. -

Topic 11: Land Plants, Part 1 (Bryophytes, Ferns & Fern Allies)
BIOL 221 – Concepts of Botany Spring 2008 Topic 11: Land Plants, part 1 (Bryophytes, Ferns & Fern Allies) A. Objectives for today’s lab . 1. Get to know 2 of the three groups of bryophytes (liverworts & mosses). 2. Get to know some of the Ferns and Fern Allies, which include some of the earliest lineages of vascular plants (represented today by Psilotum, Lycopodium, Equisetum, & Ferns—Boston (sword) fern, Staghorn fern) 3. Think about the morphological/anatomical innovations that are represented by each. Place these in the context of the origins of leaves, roots, and the fossil record and green plant phylogeny. 4. Know the general sequence of appearance: Green Algae ==> Bryophyte Lineages ==> Lycopod & Horsetail Lineages ==> Fern Lineages ==> Seed Plants (Gymnosperms & Angiosperms) B. Green Plant Phylogeny . Concepts of Botany, (page 1 of 12) C. NonVascular Free-Sporing Land Plants (Bryophytes) . C1. Liverworts . a. **LIVING MATERIAL**: Marchantia &/or Conacephalum (thallose liverworts). The conspicuous green plants are the gametophytes. With a dissecting scope, observe the polygonal outlines of the air chambers. On parts of the thallus are drier, it is easy to see the pore opening to the chamber. These are not stomata. They cannot open and close and the so the thallus can easily dry out if taking from water. Note the gemmae cups on some of that thalli. Ask your instructor what these are for. DRAW THEM TOO. b. **SLIDES**. Using the compound microscope, make observations of the vegetative and reproductive parts of various liverwort species. Note the structure of the air-chambers and the photosynthetic cells inside on the Marchantia sections! Concepts of Botany, (page 2 of 12) C2. -

Plastid Genomes of the Early Vascular Plant Genus Selaginella Have Unusual Direct Repeat Structures and Drastically Reduced Gene Numbers
International Journal of Molecular Sciences Article Plastid Genomes of the Early Vascular Plant Genus Selaginella Have Unusual Direct Repeat Structures and Drastically Reduced Gene Numbers Hyeonah Shim 1, Hyeon Ju Lee 1, Junki Lee 1,2, Hyun-Oh Lee 1,2, Jong-Hwa Kim 3, Tae-Jin Yang 1,* and Nam-Soo Kim 4,* 1 Department of Agriculture, Forestry and Bioresources, Plant Genomics & Breeding Institute, Research Institute of Agriculture and Life Sciences, College of Agriculture & Life Sciences, Seoul National University, 1 Gwanak-ro, Gwanak-gu, Seoul 08826, Korea; [email protected] (H.S.); [email protected] (H.J.L.); [email protected] (J.L.); [email protected] (H.-O.L.) 2 Phyzen Genomics Institute, Seongnam 13558, Korea 3 Department of Horticulture, Kangwon National University, Chuncheon 24341, Korea; [email protected] 4 Department of Molecular Bioscience, Kangwon National University, Chuncheon 24341, Korea * Correspondence: [email protected] (T.-J.Y.); [email protected] (N.-S.K.); Tel.: +82-2-880-4547 (T.-J.Y.); +82-33-250-6472 (N.-S.K.) Abstract: The early vascular plants in the genus Selaginella, which is the sole genus of the Selaginel- laceae family, have an important place in evolutionary history, along with ferns, as such plants are valuable resources for deciphering plant evolution. In this study, we sequenced and assembled the plastid genome (plastome) sequences of two Selaginella tamariscina individuals, as well as Se- laginella stauntoniana and Selaginella involvens. Unlike the inverted repeat (IR) structures typically found in plant plastomes, Selaginella species had direct repeat (DR) structures, which were confirmed by Oxford Nanopore long-read sequence assembly. -

Structure and Function of Spores in the Aquatic Heterosporous Fern Family Marsileaceae
Int. J. Plant Sci. 163(4):485–505. 2002. ᭧ 2002 by The University of Chicago. All rights reserved. 1058-5893/2002/16304-0001$15.00 STRUCTURE AND FUNCTION OF SPORES IN THE AQUATIC HETEROSPOROUS FERN FAMILY MARSILEACEAE Harald Schneider1 and Kathleen M. Pryer2 Department of Botany, Field Museum of Natural History, 1400 South Lake Shore Drive, Chicago, Illinois 60605-2496, U.S.A. Spores of the aquatic heterosporous fern family Marsileaceae differ markedly from spores of Salviniaceae, the only other family of heterosporous ferns and sister group to Marsileaceae, and from spores of all ho- mosporous ferns. The marsileaceous outer spore wall (perine) is modified above the aperture into a structure, the acrolamella, and the perine and acrolamella are further modified into a remarkable gelatinous layer that envelops the spore. Observations with light and scanning electron microscopy indicate that the three living marsileaceous fern genera (Marsilea, Pilularia, and Regnellidium) each have distinctive spores, particularly with regard to the perine and acrolamella. Several spore characters support a division of Marsilea into two groups. Spore character evolution is discussed in the context of developmental and possible functional aspects. The gelatinous perine layer acts as a flexible, floating organ that envelops the spores only for a short time and appears to be an adaptation of marsileaceous ferns to amphibious habitats. The gelatinous nature of the perine layer is likely the result of acidic polysaccharide components in the spore wall that have hydrogel (swelling and shrinking) properties. Megaspores floating at the water/air interface form a concave meniscus, at the center of which is the gelatinous acrolamella that encloses a “sperm lake.” This meniscus creates a vortex-like effect that serves as a trap for free-swimming sperm cells, propelling them into the sperm lake. -

Investigating Water Responsive Actuation Using the Resurrection Plant Selaginella Lepidophylla As a Model System
Investigating Water Responsive Actuation using the Resurrection Plant Selaginella lepidophylla as a Model System Véronique Brulé Department of Biology McGill University, Montréal Summer 2018 A thesis submitted to McGill University in partial fulfillment of the requirements of the degree of Doctor of Philosophy © Véronique Brulé 1 ! ABSTRACT Nature is a wealth of inspiration for biomimetic and actuating devices. These devices are, and have been, useful for advancing society, such as giving humans the capability of flight, or providing household products such as velcro. Among the many biomimetic models studied, plants are interesting because of the scope of functions and structures produced from combinations of the same basic cell wall building blocks. Hierarchical investigation of structure and composition at various length-scales has revealed unique micro and nano-scale properties leading to complex functions in plants. A better understanding of such micro and nano-scale properties will lead to the design of more complex actuating devices, including those capable of multiple functions, or those with improved functional lifespan. In this thesis, the resurrection plant Selaginella lepidophylla is explored as a new model for studying actuation. S. lepidophylla reversibly deforms at the organ, tissue, and cell wall level as a physiological response to water loss or gain, and can repeatedly deform over multiple cycles of wetting and drying. Thus, it is an excellent model for studying properties leading to reversible, hierarchical (i.e., multi length-scale) actuation. S. lepidophylla has two stem types, inner (developing) and outer (mature), that display different modes of deformation; inner stems curl into a spiral shape while outer stems curl into an arc shape. -

Megagametophyte Development of Crysanthemum Leuoanthemum L. Var. Pinnatifidum Lecoq. and Lamotte
Át1 A15MCT Q? ThE TIE$IS OF tohard ?artin for the M. 8. degree in otany Date thesis is prGented: May 14, 152 Title: ,gagarftetophyte 1evelopent of Cbrysantheuum Leuoantherniva L. var. pinnattfidw Lecoq. and L*aotte ibstraat approved: RedaCtedfOr PriVaCy Angiospern me.agarnetophytea are of three sain types; ¡onoeporic, bisporic, and tetrasortc. The nusber of weaspore nuclei taking part in seaametophyte deve1op nent te the detornining factor. In the first, only one of the tour me pore nuclei takes part in the development. in the second, two aegaspore nuclei take part in weagametophyte toration, and in the third, all tour nuclei contribute to the formation of the sature gasetophyte. In the sonosporto eight-nucleate, or Noraal type's, the seaapore aether cell nucleus undergoes the two reduottor divisions which result in the fornatton of a died and then a tetrad. The three atoropylar segasporee degenerate, and th nucleus of the one functional megaspore divides sitottoally to form a two-nucleate condition. Two further ttotic divisions fora a four-nucleate and an eight-nucleate stage. The asgagametophyte then differentiates a t)ree- celled egg apparatus, a primary endo.per oeil with two polar nuclei, and three antipodal oeils. In the tetraeporic sixteen-nucleate, or tDruea type", the nucleus of the megaepore mother cell undergoes the two reduotion divisions to form a single elongate cell oontaintn four segaspore nuolet. These then divide once attottoally to give rise to an eight-nucleate stage. )ne additional division produces a sixteen-nucleate weçagasetophyte which differentiates a three-coiled egg apparatus, a pritsary endoapers cell with to polar nuclei, arid eleven taninucleate antipodal oeils. -
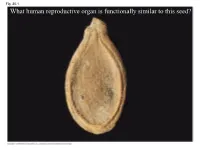
What Human Reproductive Organ Is Functionally Similar to This Seed?
Fig. 30-1 What human reproductive organ is functionally similar to this seed? Overview: Transforming the World • Seeds changed the course of plant evolution, enabling their bearers to become the dominant producers in most terrestrial ecosystems • A seed consists of an embryo and nutrients surrounded by a protective coat Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings Fig. 30-1 What human reproductive organ is functionally similar to this seed? Seeds and pollen grains are key adaptations for life on land • In addition to seeds, the following are common to all seed plants – Reduced gametophytes – Heterospory – Ovules – Pollen Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings Advantages of Reduced Gametophytes • The gametophytes of seed plants develop within the walls of spores that are retained within tissues of the parent sporophyte • Protect female gametophyte from UV and desiccation (good for the land life!) Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings Fig. 30-2 PLANT GROUP Mosses and other Ferns and other seedless Seed plants (gymnosperms and angiosperms) nonvascular plants vascular plants Reduced, independent Reduced (usually microscopic), dependent on surrounding (photosynthetic and Gametophyte Dominant sporophyte tissue for nutrition free-living) Sporophyte Reduced, dependent on Dominant Dominant gametophyte for nutrition Gymnosperm Angiosperm Sporophyte Microscopic female (2n) gametophytes (n) inside ovulate cone Microscopic Sporophyte -

Stratigraphic Distribution of Species of the Megaspore Genus Minerisporites in North America
Stratigraphic Distribution of Species of the Megaspore Genus Minerisporites in North America GEOLOGICAL SURVEY PROFESSIONAL PAPER 743-E Stratigraphic Distribution of Species of the Megaspore Genus Minerisporites in North America By ROBERT H. TSCHUDY CONTRIBUTIONS TO PALEONTOLOGY GEOLOGICAL SURVEY PROFESSIONAL PAPER 743-E Taxonomy, stratigraphic ranges, and facies significance of Minerisporites megaspores UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON: 1976 UNITED STATES DEPARTMENT OF THE INTERIOR THOMAS S. KLEPPE, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Tschudy, Robert H. Stratigraphic distribution of species of the megaspore genus Minerisporites in North America. (Contributions to Paleontology) (Geological Survey Professional Paper 743-E) Bibliography: p. Includes index. Supt. of Docs. no.: I 19.16:743-E 1. Minerisporites. 2. Paleobotany-Cretaceous. 3. Paleobotany-Paleocene. 4. Paleobotany-North America. I. Title. II. Series. III. Series: United States Geological Survey Professional Paper 743-E. QE996.T7 560' .13 76-608080 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Stock Number 024-001-02892-1 CONTENTS Page Page Abstract ............................................................................................... EI Systematics-Continued New species of Minerisporites............................................................ I Genus Minerisporites-Continued Stratigraphic distribution................................................................... I Minerisporites -

Ovule Ontogenesis and Megasporogenesis in Adesmia Latifolia (Spreng.) Vog
Revista Brasil. Bot., V.26, n.4, p.495-502, out.-dez. 2003 Ovule ontogenesis and megasporogenesis in Adesmia latifolia (Spreng.) Vog. (Leguminosae-Papilionoideae)1 MARIA C.C. MOÇO2,3 and JORGE E.A. MARIATH2 (received: January 29, 2003; accepted: September 11, 2003) ABSTRACT – (Ovule ontogenesis and megasporogenesis in Adesmia latifolia (Spreng.) Vog. (Leguminosae-Papilionoideae)). The ovule ontogenesis and the megasporogenesis events were studied under bright field, fluorescence and scanning electron microscopy. The primordium is 3-zonate and gives rise to a hemianatropous, bitegmic and crassinucellate ovule. The archesporium may consist of one or more archesporial cells, but only one undergoes meiosis, forming a linear tetrad. Normally, only a single megaspore is functional in the chalazal position, but occasionally two functional chalazal megaspores arise. The present study provides additional information on embryological characters in the Adesmieae tribe and discusses their taxonomic significance to the Leguminosae family. Key words - Adesmia, embryology, Leguminosae, megaspore, ovule RESUMO – (Ontogênese do óvulo e megasporogênese em Adesmia latifolia (Spreng.) Vog. (Leguminosae - Papilionoideae)). A ontogênese do óvulo e os eventos da megasporogênese foram estudados sob microscopia de campo claro, fluorescência e eletrônica de varredura. O primórdio é trizonado e origina um óvulo hemianátropo, bitegumentado e crassinucelado. O arquespório pode ser composto de mais de uma célula arquesporial, mas apenas uma sofre meiose formando uma tétrade linear. Normalmente, apenas um megásporo é funcional na posição calazal, mas foram registrados alguns casos em que os dois megásporos calazais eram, simultâneamente, funcionais. O presente estudo acrescenta informações sobre caracteres embriológicos na tribo Adesmieae e discute sua importância taxonômica na família Leguminosae. -

Vegetation Classification List Update for Big Bend National Park and Rio Grande National Wild and Scenic River
National Park Service U.S. Department of the Interior Natural Resource Program Center Vegetation Classification List Update for Big Bend National Park and Rio Grande National Wild and Scenic River Natural Resource Report NPS/CHDN/NRR—2011/299 ON THE COVER Chisos Basin, as viewed from Casa Grande Peak. Image provided by NPS Vegetation Classification List Update for Big Bend National Park and Rio Grande National Wild and Scenic River Natural Resource Report NPS/CHDN/NRR—2011/299 James Von Loh Cogan Technology, Inc. 8140 East Lightening View Drive Parker, Colorado 80134 Dan Cogan Cogan Technology, Inc. 21 Valley Road Galena, Illinois 61036 February 2011 U.S. Department of the Interior National Park Service Natural Resource Program Center Fort Collins, Colorado The National Park Service, Natural Resource Program Center publishes a range of reports that address natural resource topics of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Report Series is used to disseminate high-priority, current natural resource management information with managerial application. The series targets a general, diverse audience, and may contain NPS policy considerations or address sensitive issues of management applicability. All manuscripts in the series receive the appropriate level of peer review to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and designed and published in a professional manner. This report received informal peer review by subject-matter experts who were not directly involved in the collection, analysis, or reporting of the data.