Tese Doutoramento (Métodos Moleculares..).Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Presidio Phytophthora Management Recommendations
2016 Presidio Phytophthora Management Recommendations Laura Sims Presidio Phytophthora Management Recommendations (modified) Author: Laura Sims Other Contributing Authors: Christa Conforti, Tom Gordon, Nina Larssen, and Meghan Steinharter Photograph Credits: Laura Sims, Janet Klein, Richard Cobb, Everett Hansen, Thomas Jung, Thomas Cech, and Amelie Rak Editors and Additional Contributors: Christa Conforti, Alison Forrestel, Alisa Shor, Lew Stringer, Sharon Farrell, Teri Thomas, John Doyle, and Kara Mirmelstein Acknowledgements: Thanks first to Matteo Garbelotto and the University of California, Berkeley Forest Pathology and Mycology Lab for providing a ‘forest pathology home’. Many thanks to the members of the Phytophthora huddle group for useful suggestions and feedback. Many thanks to the members of the Working Group for Phytophthoras in Native Habitats for insight into the issues of Phytophthora. Many thanks to Jennifer Parke, Ted Swiecki, Kathy Kosta, Cheryl Blomquist, Susan Frankel, and M. Garbelotto for guidance. I would like to acknowledge the BMP documents on Phytophthora that proceeded this one: the Nursery Industry Best Management Practices for Phytophthora ramorum to prevent the introduction or establishment in California nursery operations, and The Safe Procurement and Production Manual. 1 Title Page: Authors and Acknowledgements Table of Contents Page Title Page 1 Table of Contents 2 Executive Summary 5 Introduction to the Phytophthora Issue 7 Phytophthora Issues Around the World 7 Phytophthora Issues in California 11 Phytophthora -

Phytophthora Occultans Man In’T Veld and Rosendahl (2015)
-- CALIFORNIA D EPAUMENT OF cdfa FOOD & AGRICULTURE ~ California Pest Rating Proposal for Phytophthora occultans Man in’t Veld and Rosendahl (2015) Current Pest Rating: Z Proposed Pest Rating: B Kingdom: Chromista, Phylum: Oomycota, Order: Peronosporales, Family: Peronosporaceae Comment Period: 11/3/2020 through 12/18/2020 Initiating Event: In June 2018, a symptomatic, mature, boxwood (Buxus microphylla) from a landscaped area inside a federal park in San Francisco County, California was submitted by a ranger to CDFA’s Meadowview Pest Diagnostics Center. Plant pathologist Suzanne Rooney-Latham isolated a Phytophthora sp. in culture and determined it to be P. occultans. As this was not considered an official sample, it was unrated. In September 2020, a sample from a block of Mexican orange blossoms (Choisya ternata) that belonged to a contractor but were slated to be planted in a new landscape at the same Federal park, was submitted to CDFA. Suzanne Rooney-Latham again isolated P. occultans. As this was an official sample and the first detection of this pathogen in California, it was given a temporary Z rating. The risk to California from P. occultans is described herein and a permanent rating is proposed. History & Status: Background: Phytophthora occultans is an oomycete pathogen that can cause a range of disease symptoms including root rot, collar, and crown rot and stem cankers on woody plants. Phytophthora spp. are widespread in temperate regions of all continents and occur in soils of natural forests, agricultural fields, landscaping, and orchards. They can persist and spread in different environments and can survive in the soil in the absence of a host. -

Phytophthora Tentaculata: a New Exotic and Invasive Disease
Phytophthora tentaculata: A New Exotic and Invasive Disease new plant pathogen in the genus pathogens of agricultural, ornamental, species and one additional genus have A Phytophthora (pronounced Fie- and forest plants. been found infected, all common in the TOF-ther-uh) has recently been found native plant nursery trade and in wild- Similar to other members of the Phy- in several California native plant nurser- lands. These include Artemisia dougla- tophthora genus, P. tentaculata releases ies and habitat restoration sites. The siana (mugwort), A. dracunculus swimming spores that move through pathogen, Phytophthora tentaculata, (tarragon), A. californica (California water and are attracted to plant root poses a risk of disease in wildlands, gar- sagebrush), Salvia species (sage), Ceano- exudates. Once infected, the pathogen dens and landscapes that use susceptible thus cuneatus (buck brush), Frangula can cause disease in susceptible plant California native and non-native plants. californica (California coffeeberry), roots. If susceptible stems are contacted, Once introduced in these areas, the Monardella villosa (coyote-mint), and infection can occur there following pathogen can generate disease for years Heteromeles arbutifolia (toyon). water movement or splash, and stem to come, potentially causing lasting envi- disease can also result from the patho- In California, P. tentaculata was first ronmental and economic impacts. gen growing into the stem from the isolated from Diplacus aurantiacus Because both native and non-native roots. Phytophthora tentaculata cannot (orange bush monkeyflower) where it California plants from nurseries can be seen with the naked eye unless grown was observed causing aboveground carry new pathogens and other pests, it in a laboratory. -

AR TICLE the Inclusion of Downy Mildews in a Multi-Locus-Dataset
doi:10.5598/imafungus.2011.02.02.07 IMA FUNGUS · VOLUME 2 · No 2: 163–171 The inclusion of downy mildews in a multi-locus-dataset and its reanalysis ARTICLE reveals a high degree of paraphyly in Phytophthora Fabian Runge1, Sabine Telle2,3, Sebastian Ploch2,3, Elizabeth Savory4, Brad Day4, Rahul Sharma2,3,5, and Marco Thines2,3,5 1University of Hohenheim, Institute of Botany 210, D-70593 Stuttgart, Germany 2Biodiversity and Climate Research Centre (BiK-F), Senckenberganlage 25, D-60325 Frankfurt am Main, Germany; corresponding author e-mail: [email protected] 3Senckenberg Gesellschaft für Naturforschung, Senckenberganlage 25, D-60325 Frankfurt am Main, Germany 4Michigan State University, Department of Plant Pathology, 104 Center for Integrated Plant Systems, East Lansing, MI 48824, USA 5Goethe University Frankfurt am Main, Department of Biological Sciences, Institute of Ecology, Evolution and Diversity, Siesmayerstr. 70, D-60323 Frankfurt (Main), Germany Abstract: Pathogens belonging to the Oomycota, a group of heterokont, fungal-like organisms, are amongst the Key words: most notorious pathogens in agriculture. In particular, the obligate biotrophic downy mildews and the hemibiotrophic AU test members of the genus Phytophthora are responsible for a huge variety of destructive diseases, including sudden downy mildews oak death caused by P. ramorum, potato late blight caused by P. infestans, cucurbit downy mildew caused by multigene phylogeny Pseudoperonospora cubensis, and grape downy mildew caused by Plasmopara viticola. About 800 species of Peronosporaceae downy mildews and roughly 100 species of Phytophthora are currently accepted, and recent studies have revealed Phytophthora that these groups are closely related. However, the degree to which Phytophthora is paraphyletic and where taxonomy exactly the downy mildews insert into this genus in relation to other clades could not be inferred with certainty to date. -
Invasive Shot Hole Borers Threatening Trees in Southern California He Polyphagous Shot Hole Borer T(PSHB) (Fig
Information for pest management professionals and pesticide applicators Green Bulletin Vol. 6 l No. 3 l December 2016 Invasive Shot Hole Borers Threatening Trees in Southern California he Polyphagous shot hole borer T(PSHB) (Fig. 1) and Kuroshio shot hole borer (KSHB) are invasive wood- boring beetles that attack dozens of tree species in Southern California, UCR ESKALEN, A. UCR ESKALEN, A. including commercial avocado groves, common landscape trees, and na- tive species in urban and wildland environments. Both beetles spread a disease called Fusarium Dieback (FD), which is caused by pathogenic fungi. Trees that are FD-susceptible may ex- perience branch dieback, canopy loss, and tree mortality (Fig. 2). Insect Vector PSHB carries three fungi: Fusarium Fig. 1. Adult female Polyphagous shot hole Fig. 2. Dead sycamore tree. euwallaceae, Graphium euwallaceae borer (Euwallaceae sp.) and Paracremonium pembeum. KSHB carries two new species of fungi: Fu- FD disease. Beetle larvae within the Internal: The fungi interrupt the sarium sp. and Graphium sp. Mature gallery in infected trees feed on the transport of water and nutrients females of both species are black and fungus, forming a symbiotic relation- in branches of affected trees, lead- 0.07 to 0.1 inches (1.8–2.5 mm) long, ship between the fungus and beetle. ing to wood discoloration which whereas males are brown and smaller Fusarium Dieback stops the flow of can vary in color from brown to than females at 0.06 inches (1.5 mm) water and nutrients in over 48 suscep- black. Shaving outer layer bark with long. The female attacks a wide variety tible tree species, which can lead to a clean knife around beetle entry of host trees forming galleries (Fig. -
A Desk Study to Review Global Knowledge on Best Practice for Oomycete Root-Rot Detection and Control
Project title: A desk study to review global knowledge on best practice for oomycete root-rot detection and control Project number: CP 126 Project leader: Dr Tim Pettitt Report: Final report, March 2015 Previous report: None Key staff: Dr G M McPherson Dr Alison Wakeham Location of project: University of Worcester Stockbridge Technology Centre Industry Representative: Russ Woodcock, Bordon Hill Nurseries Ltd, Bordon Hill, Stratford-upon-Avon, Warwickshire, CV37 9RY Date project commenced: April 2014 Date project completed April 2015 AHDB Horticulture is a Division of the Agriculture and Horticulture Development Board DISCLAIMER While the Agriculture and Horticulture Development Board seeks to ensure that the information contained within this document is accurate at the time of printing, no warranty is given in respect thereof and, to the maximum extent permitted by law the Agriculture and Horticulture Development Board accepts no liability for loss, damage or injury howsoever caused (including that caused by negligence) or suffered directly or indirectly in relation to information and opinions contained in or omitted from this document. ©Agriculture and Horticulture Development Board 2015. No part of this publication may be reproduced in any material form (including by photocopy or storage in any medium by electronic mean) or any copy or adaptation stored, published or distributed (by physical, electronic or other means) without prior permission in writing of the Agriculture and Horticulture Development Board, other than by reproduction in an unmodified form for the sole purpose of use as an information resource when the Agriculture and Horticulture Development Board or AHDB Horticulture is clearly acknowledged as the source, or in accordance with the provisions of the Copyright, Designs and Patents Act 1988. -
Phytophthora Tentaculata
Rooney-Latham, S., Blomquist, C. L., Swiecki, T., and Bernhardt, E. 2015. Phytophthora tentaculata. Forest Phytophthoras 5(1): doi: xxxxxxxx. Phytophthora tentaculata Overview Phytophthora tentaculata Kröber & Marwitz was described in 1993 in Germany on greenhouse- grown nursery ornamentals. It has since been found in Italy, Spain, China and the U.S. (California) causing a root and stem rot of many different plant species including nursery-grown native species used for habitat restoration. P. tentaculata is homothallic and is classified in Stamps group I which is characterized by the production of mostly paragynous antheridia, papillate sporangia and the production of both hyphal swellings and chlamydospores in culture. P. tentaculata is placed in phylogenetic Clade 1 (Cooke et al., 2000) with species such as P. cactorum, P. nicotianae, P. clandestina, P. iranica, P. hedraiandra and P. pseudotsugae. Etymology: refers to the spider web-like growth habit of the mycelium in culture. Figure 1. Papillate sporangia of P. tentaculata. Sporangium on the far right has an elongated neck or beak. Morphology Sporangia of P. tentaculata are spherical or ovoid to obpyriform, papillate to occasionally bipapillate and measure 10-81 x 13-52 µm (average 35.7 x 27.4 µm). They are primarily noncaducous, rarely caducous with a short pedicel, and often have elongated necks or beaks (Fig. 1). Small hyphal swellings are intercalary and often occur with hyphal branching (Fig. 2). On PARP and V8 juice agar, the hyphal growth pattern resembles a non-organized web spun by spiders in the family Theridiidae; hyphae often grow forming loops in the agar (Fig. -

Western Plant Diagnostic Network First Detector News
Western Plant Diagnostic Network1 First Detector News A Quarterly Pest Update for WPDN First Detectors Winter 2015 edition, volume 8, number 1 In this Issue Page 1: Editor’s Note Dear First Detectors, Where do insects go for the winter? Miami? Actually some insects move south for the winter, Pages 2 - 3: Insects in winter such as the famous migration of monarch butterflies to Pages 4 – 5: Phytophthora Mexico and dragon flies to warmer areas. An interview with tentaculata found in CA a specialist in insects at low temperature, Dr. Brent Sinclair, answers this question in the lead article. Two plant Pages 6 - 8: Swede Midge pathologists at the California Department of Food and Agriculture, Drs. Cheryl Blomquist and Suzanne Rooney- Page 8: NPDN online Latham, have identified a new nursery-associated Phytophthora species, P. tentaculata. A new invasive insect Pages 9 - 10: Pale cyst in Canada and the NE US, the swede midge, is a very small nematode spreads in ID and persistent pest of all Brassica crops. Cornell University Page 10: Red palm weevil has assembled a team to study the biology and control of eradicated in CA this midge: Cornell University SWEDE MIDGE Information Center for the U.S. Learn about it so you will be able to Contact us at the WPDN Regional identify the symptoms if the swede midge keeps migrating Center at UC Davis: westwards. Finally, in Pest Updates there is some bad news Phone: 530 754 2255 and some good news. Email: [email protected] Web: https://wpdn.org Editor: Richard W. Hoenisch @Copyright Regents of the -
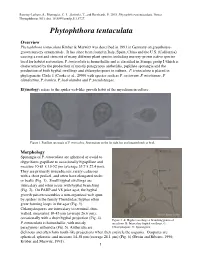
Phytophthora Tentaculata
Rooney-Latham, S., Blomquist, C. L., Swiecki, T., and Bernhardt, E. 2015. Phytophthora tentaculata. Forest Phytophthoras 5(1): doi: 10.5399/osu/fp.5.1.3727. Phytophthora tentaculata Overview Phytophthora tentaculata Kröber & Marwitz was described in 1993 in Germany on greenhouse- grown nursery ornamentals. It has since been found in Italy, Spain, China and the U.S. (California) causing a root and stem rot of many different plant species including nursery-grown native species used for habitat restoration. P. tentaculata is homothallic and is classified in Stamps group I which is characterized by the production of mostly paragynous antheridia, papillate sporangia and the production of both hyphal swellings and chlamydospores in culture. P. tentaculata is placed in phylogenetic Clade 1 (Cooke et al., 2000) with species such as P. cactorum, P. nicotianae, P. clandestina, P. iranica, P. hedraiandra and P. pseudotsugae. Etymology: refers to the spider web-like growth habit of the mycelium in culture. Figure 1. Papillate sporangia of P. tentaculata. Sporangium on the far right has an elongated neck or beak. Morphology Sporangia of P. tentaculata are spherical or ovoid to obpyriform, papillate to occasionally bipapillate and measure 10-81 x 13-52 µm (average 35.7 x 27.4 µm). They are primarily noncaducous, rarely caducous with a short pedicel, and often have elongated necks or beaks (Fig. 1). Small hyphal swellings are intercalary and often occur with hyphal branching (Fig. 2). On PARP and V8 juice agar, the hyphal growth pattern resembles a non-organized web spun by spiders in the family Theridiidae; hyphae often grow forming loops in the agar (Fig. -

R-21-63 Meeting 21-14 May 12, 2021 AGENDA ITEM 8 AGENDA ITEM
R-21-63 Meeting 21-14 May 12, 2021 AGENDA ITEM 8 AGENDA ITEM Oregon State University Phytophthora Research: Assessment of Phytophthora Soilborne Pathogens at Midpeninsula Regional Open Space District Restoration Sites GENERAL MANAGER’S RECOMMENDATIONS Receive Oregon State University’s Phytophthora Research Presentation on the Assessment of Phytophthora Soilborne Pathogens at Restoration Sites. Provide feedback on recommendations and next steps. No formal Board action necessary. SUMMARY Invasive plant pathogens in the genus Phytophthora cause significant economic and ecological damage to horticultural and agricultural industries, and native wildlands. Phytophthora species can cause negative impacts across many different plant families in a variety of native habitats when introduced into the wildlands. These species have been introduced into California wildlands via infected native plant nursery stock and through other disturbances such as soil importation. Oregon State University’s (OSU) Dr. Jennifer Parke and Dr. Ebba Peterson, have completed their research on Phytophthora in Midpeninsula Regional Open Space District (District) lands and will provide key findings and recommendations for future management actions to minimize the risk of impacts to the natural environment caused by Phytophthora. Background Soilborne Phytophthoras are a group of water molds that infect plants. There are over 150 described Phytophthora species, including the sudden oak death pathogen (Phytophthora ramorum; SOD). Although not known for certain, most experts believe that some types of Phytophthoras are native to California. They spread via spores in water, soil, or plant debris; some species are airborne. In 2004, the District began to address SOD through staff trainings, use of Best Management Practices (BMPs), updating contract language, SOD Blitzes (coordinated with the University of California, Berkeley), and by supporting research on District lands and other locations. -

Phytophthora Tentaculata
Rooney-Latham, S., Blomquist, C. L., Swiecki, T., and Bernhardt, E. 2015. Phytophthora tentaculata. Forest Phytophthoras 5(1): doi: xxxxxxxx. Phytophthora tentaculata Overview Phytophthora tentaculata Kröber & Marwitz was described in 1993 in Germany on greenhouse- grown nursery ornamentals. It has since been found in Italy, Spain, China and the U.S. (California) causing a root and stem rot of many different plant species including nursery-grown native species used for habitat restoration. P. tentaculata is homothallic and is classified in Stamps group I which is characterized by the production of mostly paragynous antheridia, papillate sporangia and the production of both hyphal swellings and chlamydospores in culture. P. tentaculata is placed in phylogenetic Clade 1 (Cooke et al., 2000) with species such as P. cactorum, P. nicotianae, P. clandestina, P. iranica, P. hedraiandra and P. pseudotsugae. Etymology: refers to the spider web-like growth habit of the mycelium in culture. Figure 1. Papillate sporangia of P. tentaculata. Sporangium on the far right has an elongated neck or beak. Morphology Sporangia of P. tentaculata are spherical or ovoid to obpyriform, papillate to occasionally bipapillate and measure 10-81 x 13-52 µm (average 35.7 x 27.4 µm). They are primarily noncaducous, rarely caducous with a short pedicel, and often have elongated necks or beaks (Fig. 1). Small hyphal swellings are intercalary and often occur with hyphal branching (Fig. 2). On PARP and V8 juice agar, the hyphal growth pattern resembles a non-organized web spun by spiders in the family Theridiidae; hyphae often grow forming loops in the agar (Fig. -

1 Diversity of Oomycetes Associated with Soybean
DIVERSITY OF OOMYCETES ASSOCIATED WITH SOYBEAN SEEDLING DISEASES By Jorge Alejandro Rojas Flechas A DISSERTATION Submitted to Michigan State University in partial fulfillment of the requirements for the degree of Plant Pathology – Doctor of Philosophy Ecology, Evolutionary Biology and Behavior – Dual Major 2016 1 ABSTRACT DIVERSITY OF OOMYCETES ASSOCIATED WITH SOYBEAN SEEDLING DISEASES By Jorge Alejandro Rojas Flechas In the United States, soybeans are produced on 76 million acres of highly productive land, but can be severely impacted by diseases caused by oomycetes. Oomycetes are part of the microbial community that is associated with plant roots and the rhizosphere, which is a dynamic and complex environment subject to the interaction of different microbes and abiotic factors that could affect the outcome of the phytobiome interaction. Depending on environmental and edaphic conditions, some oomycete species will thrive causing root and seedling rot. The identity and distribution of these pathogen species is limited. Therefore, the main questions driving my research are what oomycetes are associated with soybean seedlings and what are the roles of these species causing disease? What factors increase or reduce the impact of these species on the soybean production system? With these questions in mind, the goals of my research are: (1) to characterize the oomycete diversity associated with seedling and root rot diseases of soybean; (2) determine the role of environmental and edaphic factors on the distribution of oomycete species; (3) develop molecular diagnostic tools for Phytophthora sojae and P. sansomeana and (4) evaluate the community structure of the oomycete species associated with soybean root diseases under different conditions.