The Effect of Edaphic Factors on the Distribution and Abundance of Ants (Hymenoptera: Formicidae) in Iran
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Guide to the Ants of Sabangau
A Guide to the Ants of Sabangau The Orangutan Tropical Peatland Project November 2014 A Guide to the Ants of Sabangau All original text, layout and illustrations are by Stijn Schreven (e-mail: [email protected]), supple- mented by quotations (with permission) from taxonomic revisions or monographs by Donat Agosti, Barry Bolton, Wolfgang Dorow, Katsuyuki Eguchi, Shingo Hosoishi, John LaPolla, Bernhard Seifert and Philip Ward. The guide was edited by Mark Harrison and Nicholas Marchant. All microscopic photography is from Antbase.net and AntWeb.org, with additional images from Andrew Walmsley Photography, Erik Frank, Stijn Schreven and Thea Powell. The project was devised by Mark Harrison and Eric Perlett, developed by Eric Perlett, and coordinated in the field by Nicholas Marchant. Sample identification, taxonomic research and fieldwork was by Stijn Schreven, Eric Perlett, Benjamin Jarrett, Fransiskus Agus Harsanto, Ari Purwanto and Abdul Azis. Front cover photo: Workers of Polyrhachis (Myrma) sp., photographer: Erik Frank/ OuTrop. Back cover photo: Sabangau forest, photographer: Stijn Schreven/ OuTrop. © 2014, The Orangutan Tropical Peatland Project. All rights reserved. Email [email protected] Website www.outrop.com Citation: Schreven SJJ, Perlett E, Jarrett BJM, Harsanto FA, Purwanto A, Azis A, Marchant NC, Harrison ME (2014). A Guide to the Ants of Sabangau. The Orangutan Tropical Peatland Project, Palangka Raya, Indonesia. The views expressed in this report are those of the authors and do not necessarily represent those of OuTrop’s partners or sponsors. The Orangutan Tropical Peatland Project is registered in the UK as a non-profit organisation (Company No. 06761511) and is supported by the Orangutan Tropical Peatland Trust (UK Registered Charity No. -

Digging Deeper Into the Ecology of Subterranean Ants: Diversity and Niche Partitioning Across Two Continents
diversity Article Digging Deeper into the Ecology of Subterranean Ants: Diversity and Niche Partitioning across Two Continents Mickal Houadria * and Florian Menzel Institute of Organismic and Molecular Evolution, Johannes-Gutenberg-University Mainz, Hanns-Dieter-Hüsch-Weg 15, 55128 Mainz, Germany; [email protected] * Correspondence: [email protected] Abstract: Soil fauna is generally understudied compared to above-ground arthropods, and ants are no exception. Here, we compared a primary and a secondary forest each on two continents using four different sampling methods. Winkler sampling, pitfalls, and four types of above- and below-ground baits (dead, crushed insects; melezitose; living termites; living mealworms/grasshoppers) were applied on four plots (4 × 4 grid points) on each site. Although less diverse than Winkler samples and pitfalls, subterranean baits provided a remarkable ant community. Our baiting system provided a large dataset to systematically quantify strata and dietary specialisation in tropical rainforest ants. Compared to above-ground baits, 10–28% of the species at subterranean baits were overall more common (or unique to) below ground, indicating a fauna that was truly specialised to this stratum. Species turnover was particularly high in the primary forests, both concerning above-ground and subterranean baits and between grid points within a site. This suggests that secondary forests are more impoverished, especially concerning their subterranean fauna. Although subterranean ants rarely displayed specific preferences for a bait type, they were in general more specialised than above-ground ants; this was true for entire communities, but also for the same species if they foraged in both strata. Citation: Houadria, M.; Menzel, F. -

Fluoride Concentration of Drinking-Water of Qom, Iran
Iranian Journal of Health Sciences 2016; 4(1): 37-44 http://jhs.mazums.ac.ir Original Article Fluoride Concentration of Drinking-Water of Qom, Iran Ahmad Reza Yari 1 *Shahram Nazari 1 Amir Hossein Mahvi 2 Gharib Majidi 1, Soudabeh Alizadeh Matboo 3 Mehdi Fazlzadeh 3 1- Department of Environmental Health Engineering, School of Public Health, Qom University of Medical Sciences, Qom, Iran 2- Department of Environmental Health Engineering, School of Public Health, Tehran University of Medical Sciences, Tehran, Iran 3- Department of Environmental Health Engineering, School of Public Health, Ardabil University of Medical Sciences, Ardabil, Iran *[email protected] (Received: 4 Jul 2015; Revised: 22 Oct 2015; Accepted: 27 Dec 2015) Abstract Background and Purpose: Fluoride is a natural element essential for human nutrition due to its benefits for dental enamel. It is well-documented that standard amounts of fluoride in drinking- water can decrease the rate of dental caries. This study was conducted with the aim of measuring fluoride concentration of drinking-water supplies and urban distribution system in Qom, Iran. Materials and Methods: Results were subsequently compared against national and international standards. All sources of drinking-water of rural and urban areas were examined. To measure fluoride, the standard SPADNS method and a DR/4000s spectrophotometer were used . Results: Results showed that the mean of fluoride concentration in rural areas, mainly supplied with groundwater sources, was 0.41 mg/L, that of the urban distribution system 0.82 mg/L, that of Ali-Abad station 0.11 mg/L, and that of the private water desalination system 0.24 mg/L. -

(COVID- 19) in Iran
The epidemiological trends of coronavirus disease (COVID-19) in Iran: February 19 to March 22, 2020 Dr. Farzan Madadizadeh Research Center of Prevention and Epidemiology of Non-Communicable Disease, Department of Biostatistics and Epidemiology, School of Public Health, Shahid Sadoughi University of Medical Sciences Reyhane Sedkar ( [email protected] ) Department of Biostatistics, Shahid Beheshti University of Medical Sciences, Tehran, Iran. Research Article Keywords: Respiratory illness, Coronavirus disease 2019, Primary health care, Epidemiology Posted Date: May 18th, 2020 DOI: https://doi.org/10.21203/rs.3.rs-29367/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/17 Abstract Background The Coronavirus has crossed the geographical borders of various countries without any restrictions. This study was performed to identify the epidemiological trends of coronavirus disease (COVID-19) in Iran during February 19 to March 22, 2020. Methods This cross sectional study was carried out in 31 provinces by using the daily number of newly infected cases which was announced by the Iranian health authorities from February 19 to March 22, 2020, we explore the trend of outbreak of coronavirus disease in all provinces of Iran and determine some inuential factors such as population size, area, population density, distance from original epicenter, altitude, and human development index (HDI) for each province on its spread by Spearman correlation coecient. K-means cluster analysis (KMCA) also categorized the provinces into 10 separate groups based on CF and ACF of the infected cases at the end of the study period. (ACF). Results There were 21,638 infected, 7,913 recovered and 2,299 death cases with COVID-19 in Iran during the study period. -

Investigation of Qom Rural Area Water Network Accident in 2010 and Minimization Approaches of Accident Frequencies
•Arch Hyg Sci 2016;5(1): 33-38. RESEARCH ARTICLE •Journal Homepage: http://jhygiene.muq.ac.ir Investigation of Qom Rural Area Water Network Accident in 2010 and Minimization Approaches of Accident Frequencies Hossein Jafari Mansooriana,b, Ahmad Reza Yaric, Mohsen Ansarid, Shahram Nazarie, Mohamad Saberi Bidgolid, Gharib Majidid* aEnvironmental Health Engineering Research Center, Department of Environmental Health, Kerman University of Medical Sciences, Kerman, Iran. bYoung Researchers and Elite Club, Hamedan Branch, Islamic Azad University, Hamedan, Iran. cResearch Center for Environmental Pollutants, Qom University of Medical Sciences, Qom, Iran. dDepartment of Environmental Health Engineering, School of Public Health, Qom University of Medical Sciences, Qom, Iran. eSchool of Khalkhal Medical, Ardabil University of Medical Sciences, Ardabil, Iran. *Correspondence should be addressed to Mr. Gharib Majidi, Email: [email protected] A-R-T-I-C-L-EI-N-F-O A-B-S-T-R-A-C-T Article Notes: Background & Aims of the Study: Accidents in water networks can lead to increase the Received: Oct 25, 2015 uncounted water, costs of repair, maintenance, restoration and enter water contaminants to water network. The aim of this study is to survey the accidents of Qom rural water network Received in revised form: and choose the right approaches to reduce the number of accidents. Dec 2, 2015 Materials & Methods: In this cross-sectional study, four sector of Qom province Accepted: Dec 23, 2015 (Markazi, Dastjerd, Kahak and Qahan), were assessed over a period of 8 months (July – Available Online: Jun 1, January 2010). This study was conducted through questionnaire of Ministry of Energy. 2016 Results: The total number of accidents was 763. -

Individual Versus Collective Cognition in Social Insects
Individual versus collective cognition in social insects Ofer Feinermanᴥ, Amos Kormanˠ ᴥ Department of Physics of Complex Systems, Weizmann Institute of Science, 7610001, Rehovot, Israel. Email: [email protected] ˠ Institut de Recherche en Informatique Fondamentale (IRIF), CNRS and University Paris Diderot, 75013, Paris, France. Email: [email protected] Abstract The concerted responses of eusocial insects to environmental stimuli are often referred to as collective cognition on the level of the colony.To achieve collective cognitiona group can draw on two different sources: individual cognitionand the connectivity between individuals.Computation in neural-networks, for example,is attributedmore tosophisticated communication schemes than to the complexity of individual neurons. The case of social insects, however, can be expected to differ. This is since individual insects are cognitively capable units that are often able to process information that is directly relevant at the level of the colony.Furthermore, involved communication patterns seem difficult to implement in a group of insects since these lack clear network structure.This review discusses links between the cognition of an individual insect and that of the colony. We provide examples for collective cognition whose sources span the full spectrum between amplification of individual insect cognition and emergent group-level processes. Introduction The individuals that make up a social insect colony are so tightly knit that they are often regarded as a single super-organism(Wilson and Hölldobler, 2009). This point of view seems to go far beyond a simple metaphor(Gillooly et al., 2010)and encompasses aspects of the colony that are analogous to cell differentiation(Emerson, 1939), metabolic rates(Hou et al., 2010; Waters et al., 2010), nutrient regulation(Behmer, 2009),thermoregulation(Jones, 2004; Starks et al., 2000), gas exchange(King et al., 2015), and more. -

UNIVERSITY of KWAZULU-NATAL the Provision of Efficient Transport
UNIVERSITY OF KWAZULU-NATAL The Provision of Efficient Transport Services in the Iranian Maritime and Land Transport Interface By Farhad Razkhaneh 206525761 A dissertation submitted in fulfillment of the requirements for the degree of Doctor of Philosophy in Economics School of Accounting, Economics and Finance College of Law & Management Studies Supervisor: Professor Trevor Jones 2014 ii Acknowledgements I extend my sincere gratitude to all those who helped me through the process and preparation of this Doctoral Thesis. My sincere gratitude goes to my supervisor, Professor Trevor Jones, who meticulously read through the drafts and provided me with valuable editorial suggestions and guided me with technical comments, criticisms, guidance and support through the various stages of the writing and completion of this thesis. His efforts, knowledge and experience in international trade and transportation, ports and maritime, and merchant shipping transport related issues, have contributed towards the success of this thesis. The research and writing of this thesis, whilst at times difficult and challenging, has contributed towards my academic knowledge development, with which I hope to humbly contribute, through further writing, teaching and research, back to society. Special thanks to Professor Geoff Harris for reading earlier chapters of this thesis and providing the valuable suggestions and guidance to me. I extend my thanks to friends in the School of Accounting, Economics and Finance at the University of KwaZulu-Natal, Professor Dev Tewari and Post Doc Mr. O.B. Saiedo for their help. In addition, I am grateful for support and interest shown by colleagues in the Islamic Republic of Iran Shipping lines and individuals in the freight industry, in particular Mr. -

Borowiec Et Al-2020 Ants – Phylogeny and Classification
A Ants: Phylogeny and 1758 when the Swedish botanist Carl von Linné Classification published the tenth edition of his catalog of all plant and animal species known at the time. Marek L. Borowiec1, Corrie S. Moreau2 and Among the approximately 4,200 animals that he Christian Rabeling3 included were 17 species of ants. The succeeding 1University of Idaho, Moscow, ID, USA two and a half centuries have seen tremendous 2Departments of Entomology and Ecology & progress in the theory and practice of biological Evolutionary Biology, Cornell University, Ithaca, classification. Here we provide a summary of the NY, USA current state of phylogenetic and systematic 3Social Insect Research Group, Arizona State research on the ants. University, Tempe, AZ, USA Ants Within the Hymenoptera Tree of Ants are the most ubiquitous and ecologically Life dominant insects on the face of our Earth. This is believed to be due in large part to the cooperation Ants belong to the order Hymenoptera, which also allowed by their sociality. At the time of writing, includes wasps and bees. ▶ Eusociality, or true about 13,500 ant species are described and sociality, evolved multiple times within the named, classified into 334 genera that make up order, with ants as by far the most widespread, 17 subfamilies (Fig. 1). This diversity makes the abundant, and species-rich lineage of eusocial ants the world’s by far the most speciose group of animals. Within the Hymenoptera, ants are part eusocial insects, but ants are not only diverse in of the ▶ Aculeata, the clade in which the ovipos- terms of numbers of species. -

International Conference Integrated Control in Citrus Fruit Crops
IOBC / WPRS Working Group „Integrated Control in Citrus Fruit Crops“ International Conference on Integrated Control in Citrus Fruit Crops Proceedings of the meeting at Catania, Italy 5 – 7 November 2007 Edited by: Ferran García-Marí IOBC wprs Bulletin Bulletin OILB srop Vol. 38, 2008 The content of the contributions is in the responsibility of the authors The IOBC/WPRS Bulletin is published by the International Organization for Biological and Integrated Control of Noxious Animals and Plants, West Palearctic Regional Section (IOBC/WPRS) Le Bulletin OILB/SROP est publié par l‘Organisation Internationale de Lutte Biologique et Intégrée contre les Animaux et les Plantes Nuisibles, section Regionale Ouest Paléarctique (OILB/SROP) Copyright: IOBC/WPRS 2008 The Publication Commission of the IOBC/WPRS: Horst Bathon Luc Tirry Julius Kuehn Institute (JKI), Federal University of Gent Research Centre for Cultivated Plants Laboratory of Agrozoology Institute for Biological Control Department of Crop Protection Heinrichstr. 243 Coupure Links 653 D-64287 Darmstadt (Germany) B-9000 Gent (Belgium) Tel +49 6151 407-225, Fax +49 6151 407-290 Tel +32-9-2646152, Fax +32-9-2646239 e-mail: [email protected] e-mail: [email protected] Address General Secretariat: Dr. Philippe C. Nicot INRA – Unité de Pathologie Végétale Domaine St Maurice - B.P. 94 F-84143 Montfavet Cedex (France) ISBN 978-92-9067-212-8 http://www.iobc-wprs.org Organizing Committee of the International Conference on Integrated Control in Citrus Fruit Crops Catania, Italy 5 – 7 November, 2007 Gaetano Siscaro1 Lucia Zappalà1 Giovanna Tropea Garzia1 Gaetana Mazzeo1 Pompeo Suma1 Carmelo Rapisarda1 Agatino Russo1 Giuseppe Cocuzza1 Ernesto Raciti2 Filadelfo Conti2 Giancarlo Perrotta2 1Dipartimento di Scienze e tecnologie Fitosanitarie Università degli Studi di Catania 2Regione Siciliana Assessorato Agricoltura e Foreste Servizi alla Sviluppo Integrated Control in Citrus Fruit Crops IOBC/wprs Bulletin Vol. -

The First Record Among Dolichoderinae (Formicidae) of Parasitism by Strepsiptera
Insectes Soc. 50 (2003) 148–150 0020-1812/03/020148-03 Insectes Sociaux DOI 10.1007/s00040-003-0637-9 © Birkhäuser Verlag, Basel, 2003 Research article The first record among Dolichoderinae (Formicidae) of parasitism by Strepsiptera D.P. Hughes 1,G.Moya-Raygoza 2 and J. Kathirithamby 1 1 Department of Zoology, University of Oxford, South Parks Road, OX1 3PS, UK, e-mail:[email protected] 2 Departamento de Botanica y Zoologia, C.U.C.B.A., Universidad de Guadalajara, Km 15.5 carretera Guadalajara-Nogales, Las Agujas, Zapopan, C.P. 45110, Apdo, Postal 139, Jalisco, Mexico Received 7 November 2002; revised 19 December 2002; accepted 2 January 2003. Summary. We present the first record of parasitism of Host location is conducted by the 1st instar larvae of the par- Dolichoderus bispinosus nests by Strepsiptera belonging to asite, which enter the colony, likely to be via phoretic trans- the family Myrmecolacidae. This becomes only the four- port, where they are assumed to infect larval ants. Unlike teenth species of ant and the fifth subfamily to be identified other endoparasitoids of ants, such as eucharitids (Hymen- as a host to Strepsiptera. Of the three colonies examined all optera: Eucharitidae), Strepsiptera remain associated with were parasitized. Prevalence of parasitism among adult ants the host even after host metamorphosis. Following host eclo- was less than 2% in each case. However, among alate males sion the male parasite forms a pupa, the anterior portion, i.e. of one colony, nearly 24% were parasitized. In conjunction cephalothecae, of which extrudes through the intersegmental with a reanalysis of previously published data we discuss the membranes of the gaster. -
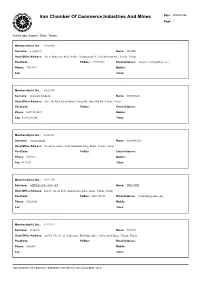
Iran Chamber of Commerce,Industries and Mines Date : 2008/01/26 Page: 1
Iran Chamber Of Commerce,Industries And Mines Date : 2008/01/26 Page: 1 Activity type: Exports , State : Tehran Membership Id. No.: 11020060 Surname: LAHOUTI Name: MEHDI Head Office Address: .No. 4, Badamchi Alley, Before Galoubandak, W. 15th Khordad Ave, Tehran, Tehran PostCode: PoBox: 1191755161 Email Address: [email protected] Phone: 55623672 Mobile: Fax: Telex: Membership Id. No.: 11020741 Surname: DASHTI DARIAN Name: MORTEZA Head Office Address: .No. 114, After Sepid Morgh, Vavan Rd., Qom Old Rd, Tehran, Tehran PostCode: PoBox: Email Address: Phone: 0229-2545671 Mobile: Fax: 0229-2546246 Telex: Membership Id. No.: 11021019 Surname: JOURABCHI Name: MAHMOUD Head Office Address: No. 64-65, Saray-e-Park, Kababiha Alley, Bazar, Tehran, Tehran PostCode: PoBox: Email Address: Phone: 5639291 Mobile: Fax: 5611821 Telex: Membership Id. No.: 11021259 Surname: MEHRDADI GARGARI Name: EBRAHIM Head Office Address: 2nd Fl., No. 62 & 63, Rohani Now Sarai, Bazar, Tehran, Tehran PostCode: PoBox: 14611/15768 Email Address: [email protected] Phone: 55633085 Mobile: Fax: Telex: Membership Id. No.: 11022224 Surname: ZARAY Name: JAVAD Head Office Address: .2nd Fl., No. 20 , 21, Park Sarai., Kababiha Alley., Abbas Abad Bazar, Tehran, Tehran PostCode: PoBox: Email Address: Phone: 5602486 Mobile: Fax: Telex: Iran Chamber Of Commerce,Industries And Mines Center (Computer Unit) Iran Chamber Of Commerce,Industries And Mines Date : 2008/01/26 Page: 2 Activity type: Exports , State : Tehran Membership Id. No.: 11023291 Surname: SABBER Name: AHMAD Head Office Address: No. 56 , Beside Saray-e-Khorram, Abbasabad Bazaar, Tehran, Tehran PostCode: PoBox: Email Address: Phone: 5631373 Mobile: Fax: Telex: Membership Id. No.: 11023731 Surname: HOSSEINJANI Name: EBRAHIM Head Office Address: .No. -

Review of Ants (Hymenoptera: Formicidae) from Jordan
ANNALS OF THE UPPER SILESIAN MUSEUM IN BYTOM ENTOMOLOGY Vol. 29 (online 002): 1–26 ISSN 0867-1966, eISSN 2544-039X (online) Bytom, 30.03.2020 Lech Borowiec1 , Sebastian Salata2 Review of ants (Hymenoptera: Formicidae) from Jordan http://doi.org/10.5281/zenodo.3733156 1 Department of Biodiversity and Evolutionary Taxonomy, University of Wrocław, Przybyszewskiego 65, 51-148 Wrocław, Poland, e-mail: [email protected] 2 Department of Entomology, California Academy of Sciences, San Francisco, CA 94118, USA, e-mail: [email protected] Abstract: We present an updated checklist of ant species known from Jordan. In total we list 58 species and 26 morphospecies identified to genus or species group level. Ten species are recorded from the country for the first time: Aphaenogaster schmitzi Forel, 1910, Camponotus gestroi Emery, 1878, Camponotus rebeccae Forel, 1913, Crematogaster warburgi Menozzi, 1933, Hypoponera punctatissima (Roger, 1859), Lepisiota bipartita (Smith, 1861), Monomorium luteum Emery, 1881, Monomorium venustum (Smith, 1858), Tapinoma simrothi Krausse, 1911, and Trichomyrmex destructor (Jerdon, 1851). We also recognize 26 morphospecies which determination, due to lack of comprehensive taxonomic studies, is unachievable and some of them can represent species new to science. Furthermore, we list doubtful records of ten taxa: Camponotus aethiops (Latreille, 1798), Cataglyphis bicolor (Fabricius, 1793), Cataglyphis livida (André, 1881), Messor concolor Santschi, 1927, Messor meridionalis (André, 1883), Plagiolepis pallescens maura Santschi, 1920, Tapinoma erraticum (Latreille, 1798), Tapinoma nigerrimum (Nylander, 1856), Temnothorax luteus (Forel, 1874), and Tetramorium caespitum (Linnaeus, 1758), and discuss their possible affiliation with species of documented and certain presence in Jordan. Key words: ants, biogeography, new records, the Middle East.